



Developmental Neurobiology Unit
Principal Investigator: Ichiro Masai
Research Theme: Mechanisms underlying neuronal differentiation and degeneration in the zebrafish retina

Abstract
The vertebrate neural retina is derived from the ventral region of the forebrain. In the neural retina, six classes of retinal neurons, including photoreceptors, differentiate and form the neural circuit that mediates visual transduction. Thus, the retina provides a good model for studying cell differentiation in the vertebrate brain. Furthermore, more than a hundred hereditary retinal diseases causing photoreceptor degeneration have been identified in humans. Understanding the pathological processes of photoreceptor degeneration is medically important. We are currently investigating the mechanisms underlying retinal cell differentiation and photoreceptor degeneration, using zebrafish as an animal model. We mainly focus on two research topics: neurogenesis and photoreceptor degeneration. In the project on retinal neurogenesis, we have shown that signaling molecules from at least five protein families – the Hedgehog, Fibroblast growth factor, Wnt, Notch and Histone deacetylase families – regulate the spatial and temporal pattern of retinal neurogenesis in zebrafish. We are identifying regulatory factors involved in the Histone deacetylase 1-dependent regulation of retinal neurogenesis. In the project on photoreceptor degeneration, we have identified zebrafish mutants showing photoreceptor degeneration. By cloning genes whose mutations cause photoreceptor degeneration, we are investigating the mechanisms underlying the pathological processes of photoreceptor degenerations in these mutants.1. Staff
- Dr. Yuko Nishiwaki, Researcher
- Dr. Toshiaki Mochizuki, Researcher
- Dr. Fumiyasu Imai (departed June 2010), Researcher
- Dr. Yukihiro Yoshimura, Researcher
- Dr. Ekramul Islam (started July 2010), Researcher
- Ms. Asuka Yoshizawa, Technical Staff
- Dr. Shohei Suzuki, Technical Staff
- Mr. Yutaka Kojima, Technical Staff
- Ms. Eri Oguri (Temporary), Technical Staff
- Ms. Yayoi Tomoyose, Research Assistant
- Ms. Yuki Tamae, Research Assistant
- Ms. Ayako Nameki (started December 2010), Research Assistant
- Ms. Ayako Gima, Research Administrator/Secretary
2. Collaborations
Nothing to report
3. Activities and Findings
3.1 Mechanisms regulating retinal neurogenesis in zebrafish
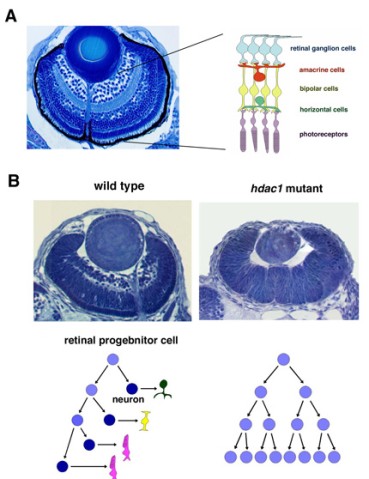
Figure 1: (A) Three dpf zebrafish retina. Six major classes of retinal neurons differentiate and form layered structures. (B) Three dpf wild-type and hdac1 mutant retinas. In the hdac1 mutant, retinal progenitor cells fail to exit from the cell cycle and continue to proliferate like a cancer, suggesting that HDAC1 regulate the switch from proliferation to differentiation in the zebrafish retina.
The vertebrate retina is initially specified in the anterior neural plate and evaginates from the ventral region of the forebrain as the optic cup. In this region, six major classes of retinal neurons and one class of glial cells differentiate to form the neural circuit that mediates phototransduction and visual processing (Fig. 1A). Thus, the retina provides an excellent model for studying the mechanisms underlying cell differentiation and neural circuit formation in the developing brain. In the developing zebrafish retina, neurogenesis is initiated at a few cells adjacent to the optic stalk and progresses to the entire neural retina. This pattern of retinal neurogenesis provides a good model for studying the mechanism regulating the spatial and temporal pattern of neurogenesis in the nervous system. Previous studies, including ours, suggest that at least five families of signaling molecule regulate the pattern of retinal neurogenesis in zebrafish (Fig. 2). Fibroblast growth factors (Fgfs) are expressed in the optic stalk and are required for the initial induction of retinal neurogenesis in zebrafish. The Hedgehog (Hh) signaling pathway is important for the progression of retinal neurogenesis in zebrafish. Activation of the Wnt signaling pathway promotes cell proliferation in the zebrafish retina, and activation of the Notch pathway inhibits retinal neurogenesis. Previously, we identified a zebrafish histone deacetylase 1 (hadc1) mutant, in which retinal progenitor cells fail to exit from the cell cycle and continue to proliferate (Fig. 1B). HDAC1 promotes retinal neurogenesis by suppressing both the Wnt and Notch signaling pathways. HDAC1 is recruited to several transcription repressor and corepressor complexes as well as those in the Wnt and Notch pathways. To elucidate the HDAC1-dependent regulation of retinal neurogenesis, we have identified a new mutation, oki150, that modifies the neurogenic defects in heterozygous hdac1 mutants. We are currently cloning the gene containing the oki150 mutation.

Figure 2: Molecular network regulating retinal neuroegnesis in zebrafish. FGF and Hh regulate the induction and progression of retinal neurogenesis in zebrafish. Wnt and Notch signaling pathways promote cell proliferation and neurogenic inhibition, respectively. HDAC1 promotes retinal neurogenesis by suppressing both Wnt and Notch signaling pathways. A novel mutation namely oki150 may interact with HDAC1 to promote neurogenesis.
3.2 Roles of apicobasal cell polarity in retinal neurogenesis
Cell and epithelial polarity is compromised in various human cancers. The loss of apicobasal cell polarity and the disruption of adherens junctions induce hyperplasia in the mouse developing brain and Drosophila imaginal discs. However, the relationship between the loss of cell polarity, proliferation, and differentiation has been discussed for a long time. Three groups of proteins are central in the establishment and maintenance of apicobasal cell polarity. The Crumbs–Stardust and Par3–Par6–aPKC protein complexes promote apical-membrane domain identity, whereas the Lethal giant larvae–Scribble–Discs large protein complex promotes basolateral membrane identity. The apical junctional complex separates the apical and basolateral membrane domains and links internal cell polarity with three-dimensional tissue organization. We examined the role of cell polarity and epithelial integrity in retinal neurogenesis. We focused on two zebrafish mutants, n-cadherin (ncad) and nagie oko (nok), in both of which apicobasal cell polarity and adherens junctions are disrupted in the retinal epithelium. We found that the fraction of proliferating cells increased in these mutant retinas during neurogenic stages. We examined the mode of cell division during neurogenesis and found that the switch from proliferative cell division to neurogenic cell division is compromised in these mutant retinas, resulting in the reduction of retinal neurogenesis. Furthermore, Notch and Hh–protein kinase A (PKA) signaling pathways are involved in the neurogenic defects in these mutant retinas. These findings suggest that the loss of apicobasal cell polarity affect the mode of cell division during retinal neurogenesis in zebrafish through the Notch and Hh–PKA signaling pathways. Cell polarity regulators function as neurogenic promoting factors in the zebrafish retina. These findings were published this fiscal year.
3.3 Mechanisms regulating neuronal apoptosis in the zebrafish retina
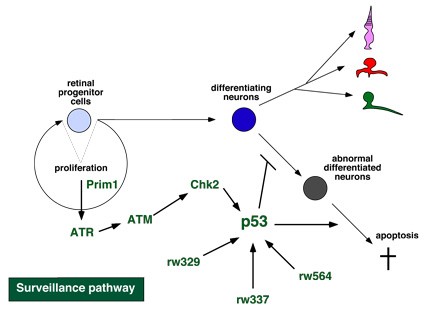
Figure 3: Surveillance pathway in the developing zebrafish retina. Prim1 activates DNA replication checkpoint ATR. ATR activates the DNA damage response, ATM and Chk2. Chk2 activate p53, which repair DNA damages or induce apoptosis to eliminate abnormal differentiated cells.
Apoptosis is observed in a developing retina and is believed to remove abnormal differentiated cells. However, it is unknown how the apoptotic pathway is regulated during neurogenesis. Previously, we identified four zebrafish mutants that show severe apoptosis of differentiating retinal neurons. Previously, we characterized one of these apoptotic mutants, pinball eye (piy). The piy mutant gene encodes the small subunit of DNA primase (Prim1). Prim1 is essential for DNA replication and is also required for the activation of the DNA damage response. We found that apoptosis in the piy mutant depends on the DNA damage response pathway, which activates a series of signaling molecules, Ataxia telangiectasia mutated (ATM), Checkpoint kinase 2 (Chk2), and the tumor suppressor p53. These findings suggest that the DNA damage response and its downstream target p53 determine whether retinal cells will differentiate normally or undergo apoptosis during retinal neurogenesis (Fig. 3). In addition to the piy mutant, retinal apoptosis in three other apoptotic mutants, rw329, rw337, and rw564, depends on p53. To elucidate the p53-dependent molecular mechanism that determines whether retinal cells are eliminated by apoptosis, we are cloning these mutant genes. Recently, we identified the rw564 gene, which encodes a novel protein containing WD40 repeats, and are characterizing its function.
3.4 Systematic understanding of the growth of the vertebrate eye
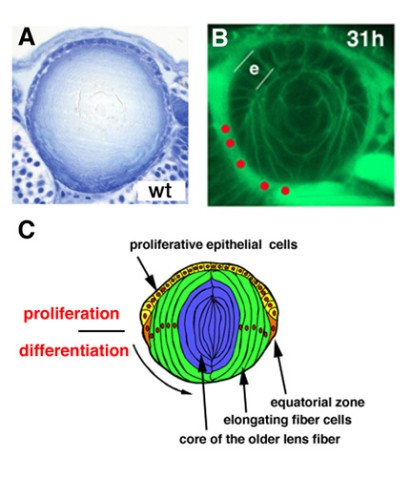
Figure 4: Lens development. (A) Zebrafish wild-type lens at 7dpf. (B) Labeling of 36 hpf wild-type lens with fluorescent lipid (green). Position of basal foots of differentiating lens fiber cells is indicated by red circles. (C) Schematic drawing of lens structures. Anterior lens epithelial cells (yellow) are proliferative and start to differentiate into lens fiber cells (green) at the equator. Differentiating lens fiber cells elongate bilaterally to cover the old lens fiber core (blue).
The lens is an intraocular tissue that supports vision. During development, the lens delaminates from the epidermal ectoderm and forms the lens vesicle. In the lens vesicle, the posterior epithelial cells differentiate into primary lens fiber cells, whereas the anterior epithelial cells are maintained as proliferating lens epithelium. At the boundary between the anterior and posterior epithelium, called the equator, epithelial cells start to differentiate into secondary lens fiber cells, which elongate and cover the old lens fiber core in an onion-like manner (Fig. 4). The growth rate of the lens must correlate with that of the retina, because visual images need to be focused exactly on the photoreceptor layer through lens-mediated refraction. However, the mechanism that coordinates the growth rates of the retina and the lens is unknown.
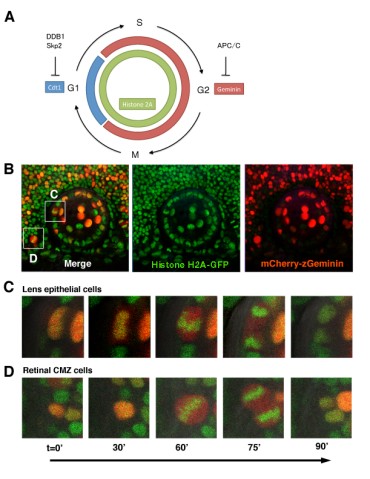
Figure 5: Visualization of cell-cycle progression in the zebrafish retina and lens. (A) Zebrafish Cecyil transgenic system. Cdt and Geminin expression is regulated through protein degradation by E3 ubiqutin ligase DDB1/Skp2 and APC/C, respectively. Cdt (Blue) and Geminin (red) are expressed in G1 and S/G2/M phase, respectively. GFP-tagged Histone2A transgene (green) labels nuclei of all retinal and lens cells. (B) Confocal images of zebrafish transgenic line carrying GFP-tagged Histone 2A (green) and mCherry-tagged Geminin (red). (C and D) Confocal images of retinal (C) and lens (D) M-phase cells in the zebrafish transgenic line carrying GFP-tagged Histone 2A (green) and mCherry-tagged Geminin (red). In the mitosis, GFP-tagged Histone2A expression (green) becomes condensed in segregating chromosomes and mCherry-Geminin expression (red) is the highest. Alignment of condensed chromosomes and segregating direction are clearly visualized.
We currently take two approaches: first, in vivo imaging of cell behaviors such as proliferation, differentiation and cell movement in the zebrafish lens epithelium, and second, analysis of zebrafish mutants showing defects in lens proliferation and differentiation. In the first project on in vivo imaging of cell behaviors in the lens, we use the zebrafish Cecyil transgenic strain, in which cell-cycle phases can be differentially visualized by fluorescent proteins (Sugiyama et al., 2009, Proc Natl Acad Sci USA 106, 20812–20817). The cell cycle consists of the G1, S, G2, and M phases. Two cell-cycle regulators, Cdt1 and Geminin, are expressed in the G1 and the S-G2-M phase, respectively. In the Cecyil transgenic line, the cell-cycle-dependent expression of Cdt1 and Geminin is illuminated with different fluorescent proteins (Fig. 5A). In zebrafish carrying genes encoding mCherry-tagged Geminin and green fluorescent protein (GFP)-tagged Histone 2A, we can clearly distinguish three different phases, G1, S-G2, and M phase, in the developing retina and lens (Fig. 5B-D). We have observed the pattern of cell-cycle progression, cell division, and cell movement in the zebrafish lens from 36 hours post fertilization (hpf) to 72 hpf using time-lapse imaging. In the developing lens, we found that rapidly proliferating cells are positioned in a limited zone just anterior to the equator in the lens epithelium. This observation is consistent with previous reports in the lenses of other vertebrates.
In the second approach, we have identified zebrafish mutants showing defects in lens proliferation and lens fiber cell differentiation. Recently, we characterized the zebrafish psmd6 mutant, in which proliferation in the lens epithelium and lens fiber cell differentiation are compromised. Psmd6 is a regulatory subunit of the proteasome. The ubiquitin and proteasome system (UPS) is important for the degradation of unnecessary proteins. Furthermore, the UPS regulates various cellular and developmental processes by degrading their regulatory proteins. These data suggest that UPS-mediated protein degradation is important for cell proliferation in the lens epithelium and in lens fiber differentiation in zebrafish. The findings on the zebrafish psmd6 mutant were published this fiscal year.
3.5 Mechanisms underlying photoreceptor degeneration
Photoreceptors are highly specialized neurons that efficiently detect light stimuli. The phototransduction proteins are located within a modified and greatly enlarged cilium, termed the outer segment (OS), which protrudes from the apical face of each photoreceptor (Fig. 6A). The OS is filled with flattened membrane sacs called discs and is constantly renewed by synthesis and assembly at its base and by shedding of old material from the tip. However, the mechanisms underlying the establishment of such sophisticated photoreceptor ultrastructures remain to be elucidated. To date, more than a hundred genes associated with inherited photoreceptor degeneration have been identified in humans (see the homepage of the Retinal Information Network at http://www.sph.uth.tmc.edu/Retnet/). Photoreceptor degeneration is unique in that mutations in genes encoding proteins of great functional diversity lead to the death of cells, for example proteins involved in phototransduction, retinol metabolism and the transport of proteins to the photoreceptive membrane through intraflagellar transport. However, the mechanisms underlying pathological processes in photoreceptor degeneration also remain to be elucidated.
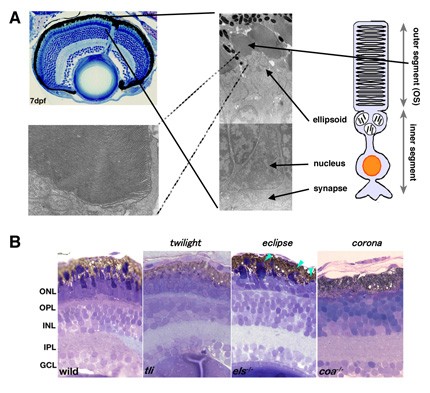
Figure 6: Photoreceptor structures and zebrafish mutant showing photoreceptor degeneration. (A) 7dpf zebrafish retina (right top panel) and electron microscopic images of photoreceptor cells (left panel) and the outer segment (right bottom panel). Photoreceptors show specialized structures: the outer segment (OS) and the inner segment consisting of mitochondria-rich ellipsoid, nucleus and synapse. In the OS, photoreceptive membranes are multi-stacked. (B) Semi-thin sections of 7dpf zebrafish photoreceptor degeneration mutants: twilight (tli), eclipse (els) and corona (coa). In the tli mutant, photoreceptors lack the OS. In the els mutant, photoreceptor cell shape is abnormal. In the coa mutant, almost all photoreceptors are absent. ONL, the outer nuclear layer; OPL, the outer plexiform layer; INL the inner nuclear layer; IPL, the inner plexiform layer; GCL, retinal ganglion cell layer.
To determine the mechanisms involved in establishing the structural integrity of photoreceptors and in photoreceptor degeneration, we have screened zebrafish mutants with disrupted visual behaviors by testing the optokinetic response. We identified three zebrafish mutants, twilight (tli), eclipse (els), and corona (coa), in which photoreceptors differentiate but later degenerate (Fig. 6B). Previously, we cloned the els mutant gene and found that it encodes the cone-specific subunit of cGMP-phosphodiesterase 6 (PDE6c). PDE6c mediates phototransduction in vertebrate cone photoreceptors. Indeed, only cone-specific phototransduction is disrupted in the els mutant. In human genetic diseases associated with photoreceptor degeneration, mutations in rod-specific subunits of PDE6 have been reported. However, no mutations in the PDE6c gene have been found in humans. We found that cones are selectively eliminated but rods are retained at the adult stage in the els mutant. These observations suggest that PDE6c is important not only for phototransduction but also for survival of cone photoreceptors. In human and mouse mutations in rod-specific subunits of PDE6, the degeneration of cones follows that of rods. The specific loss of cones in the els mutant suggests a non-reciprocal interdependence between rod and cone survival. Recently, it has been reported that mutations in the human PDE6C gene cause progressive cone dystrophy and achromatopsia, suggesting great similarity between humans and zebrafish. The els mutant will shed light on our understanding of the pathological processes underlying this human cone photoreceptor disorder. To understand the mechanism underlying photoreceptor degeneration more deeply, we are currently cloning another two mutant genes, tli and coa.
4. Publications
4.1 Journals
Imai, F. Yoshizawa, A. Fujimori-Tonou, N. Kawakami, K. Masai, I. The ubiquitin proteasome system is required for cell proliferation of the lens epithelium and for differentiation of lens fiber cells in zebrafish. Development 137, 3257-69 (2010).
Yamaguchi, M. Imai, F. Tonou-Fujimori, N. Masai, I. Mutations in N-cadherin and a Stardust homolog, Nagie oko, affect cell-cycle exit in zebrafish retina. Mech Dev 127, 247-64 (2010).
Ohata, S. Aoki, R. Kinoshita, S. Yamaguchi, M. Tsuruoka-Kinoshita, S. Tanaka, H. Wada, H. Watabe, S. Tsuboi, T. Masai, I. Okamoto, H. Dual Roles of Notch in Regulation of Apically Restricted Mitosis and Apicobasal Polarity of Neuroepithelial Cells. Neuron 69, 215-30 (2011).
4.2 Books and other one-time publications
Nothing to report.4.3 Oral and Poster Presentations
Yoshimura, Y. Oguri, E. Hanahara, N. Fujimori-Tonou, N. Masai, I. A novel WD40-repeat containing protein required for recruitment of XPC to DNA damaged sites in nucleotide excision repair. 9th International conference on zebrafish development and genetics, University of Wisconsin-Madison, Madiosn, Wisconsin, USA, 16-20 June 2010.
Mochizuki, T. Masai, I. Zebrafish VPS45 is essential for lens epithelial integrity and lens fiber differentiation. 9th International conference on zebrafish development and genetics, University of Wisconsin-Madison, Madiosn, Wisconsin, USA, 16-20 June 2010.
Ohata, S., Aoki, R., Kinoshita, S., Yamaguchi, M., Tsuruoka-Kinoshita, S., Tanaka, H., Wada, H., Tsuboi, T., Masai, I., Okamoto, H. Dual roles of Notch in regulation of apically restricted mitosis and apicobasal polarity neuroepithelial cells. The 43rd Annual meeting of Japanese scoeity of developmental biologists, Kyoto, Japan, 20-23 June 2010.
Imai, F., Yoshizawa, A., Masai, I. Ubiquitin poteasome system is essential for the lens development. The 43rd Annual meeting of Japanese scoeity of developmental biologists, Kyoto, Japan, 20-23 June 2010.
Mochizuki, T. Roles of Vps45 in lens cell differentiation and morphogenesis. The 3rd Retina research meeting, Tokyo, Japan, 4 Dec 2010. (in Japanese)
Masai, I. A novel WD40 repeat protein mediates reapair and inhibits apoptosis in retinal cells. The 3rd Retina research meeting, Tokyo, Japan, 4 Dec 2010. (in Japanese)
Mochizuki, T. Masai, I. Zebrafish VPS45 is essential for lens epithelial integrity and lens fiber differentiation. BMB2010 (Joint meeting of the 33rd annual meeting of Japanese society of Molecular Biology and the 83rd annual meeting of Japanese society of Biochemistry, Kobe, Japan, 7-10 Dec 2010. (in Japanese)
Yoshimura, Y. Oguri, E. Hanahara, N. Fujimori-Tonou, N. Masai, I. A novel WD40-repeat containing protein required for recruitment of XPC to DNA damaged sites in nucleotide excision repair. BMB2010 (Joint meeting of the 33rd annual meeting of Japanese society of Molecular Biology and the 83rd annual meeting of Japanese society of Biochemistry, Kobe, Japan, 7-10 Dec 2010. (in Japanese)
5. Intellectual Property Rights and Other Specific Achievements
Nothing to report.6. Meetings and Events
6.1 OIST Course
- Title: OIST Development Neurobiology Course
- Date: 12-23 July 2010
- Venue: OIST Seaside House
- Co-organizers:
- Mary-Ann Price (OIST)
- David L. van Vactor (Harvard Medical School, OIST)
- Robert Baughman (OIST)
- Akinao Nose (University of Tokyo)
- Speakers:
- Yoshiki Sasai (RIKEN CDB, Japan)
- Ichiro Masai (OIST)
- Li-Huei Tsai (Massachusetts Institute of Technology)
- Mary-Ann Price (OIST)
- David van Vactor (Harvard Medical School, OIST)
- Masayuki Miura (University of Tokyo)
- Frank Gertler (Massachusetts Institute of Technology)
- Elke Stein (Yale University)
- Akinao Nose (University of Tokyo)
- Tadashi Uemura (Kyoto University)
- Kang Shen (Stanford University)
- Morgan Sheng (Genentech Inc)
- Hitoshi Okamoto (RIKEN BSI)
- John Flanagan (Harvard Medical School)
- Nirao M. Shah (UCSF)
- Hideyuki Okano (Keio University)
- Alexander J. Whitworth (University of Sheffield)
- Thomas Walz (Harvard Medical School)
6.2 OIST Seminar
- Title: Mechanics of epithelial morphogenesis in Drosophila
- Date: 29 July 2010
- Venue: OIST Campus
- Speaker: Dr. Shigeo Hayashi (RIKEN, CDB, Japan)
6.3 OIST Seminar
- Title: Essential roles of PTEN/PI3K signaling in proper retinal network formation
- Date: 22 December 2010
- Venue: OIST Campus
- Speaker: Dr. Kiyo Sakagami (Jules Stein Eye Institute, University of California Los Angeles)
6.4 OIST Seminar
- Title: An axonal switch required for establishing neural circuits
- Date: 10 March 2010
- Venue: OIST Campus
- Speaker: Dr. Junichi Kawada (Department of Neurology, Northwestern University Feinberg School of Medicine, USA)
7. Others
7.1 Conference
- Title: 9th International Conference of Zebrafish Development and Genetics
- Date: 16-20 June 2010
- Venue: University of Wisconsin, Madison
- Co-organizers:
- Michael Granato (University of Pennsylvania)
- Corinne Houart (King’s College London)
- Cecilia Moens (Fred Hutchinson Cancer Research Center)
- Derek Stemple (Wellcome Trust Sanger Institute)
- William Talbot (Stanford University)
- Remarks: This conference is the largest international conference in zebrafish research community.