



Neurobiology Research Unit
Principal Investigator: Jeff Wickens
Research Theme: Cellular mechanisms of learning and adaptive behaviour

Abstract
The overall aim of the Neurobiology Research Unit is to elucidate the cellular mechanisms of learning and adaptive behaviour in the brain. We focus on the mammalian basal ganglia, a set of brain structures implicated in reward and intentional action. Previously we found that reward causes physical changes in the neural connections of the basal ganglia, which are associated with learning. Now we aim to discover the mechanisms that govern these changes in neural connections, concentrating on the actions of nenurochemicals - such as dopamine - that are released by rewards. Research activity has focused on experimental studies of synaptic plasticity in the corticostriatal pathway, and theoretical studies of striatal network dynamics and reinforcement mechanisms important in learning. Our hypothesis is that molecular signaling networks activated by dopamine define precise rules for activity-dependent synaptic plasticity. We are investigating synaptic plasticity in live brain tissue using electrophysiological, molecular and computational approaches at multiple levels of analysis. We are undertaking studies using 2-photon microscopy to measure events taking place at individual dendritic spines. Conventional patch-clamp recording is being used to measure synaptic plasticity. Lines of transgenic mice have been established in which different types of cells can be definitively identified. Using these approaches we are investigating (i) precise timing requirements for synaptic plasticity (ii) cell-type specific differences in synaptic plasticity, and (iii) dendritic mechanisms underlying synaptic plasticiy. The long-term aims of these experiments are to determine rules for induction of synaptic plasticity and to elucidate their underlying cellular mechanisms. We are also undertaking theoretical studies at cellular, network and systems levels, which aim to integrate the experimental findings with network level activity and examine their implications for understanding and treating disorders.
1. Staff
- Dr. Tomomi Shindou, Researcher
- Dr. Catherine Vickers, Researcher
- Dr. Mayumi Ochi-Shindou, Researcher
- Dr. Adam Ponzi, Researcher
- Dr. Luca Aquili, Researcher
- Dr. Takashi Nakano, Researcher
- Dr. Saori Miura, Technical Staff
- Mr. Andy Liu, Technical Staff
- Mr. Kavinda Liyanagama, Technical Staff
- Mr. Yu-Ting Li, Graduate Student
- Ms. Yi-Ling Huang, Research Assistant
- Ms. Yukako Suzuki, Research Administrator
2. Collaborations
- Theme: Cellular and Behavioural Mechanisms of Hyperactivity and Movement Disorders
- University of Otago, New Zealand
- Type of collaboration: Joint research
- Researchers: Dr. Brian Hyland, Dr. Brent Alsop, Dr. Alison Mercer, Dr. Gail Tripp
- Theme: Research on neuroplasticity using wireless dopamine sensing and microPET
- Institute of Biomedical Engineering, National Cheng Kung Univerity, Taiwan
- Type of collaboration: Research Collaboration
- Researchers: Mr. Yu-Ting Li
3. Activities and Findings
The Neurobiology Research Unit was established January 29, 2007. During the first year of operation, the laboratory interior was completed and the equipment was commissioned. recruitment has continued and in the past year the Unit grew to its final size when two new researchers joined us. Research activity has focused on experimental studies of synaptic plasticity in the corticostriatal pathway, and theoretical studies of striatal network dynamics and reinforcement mechanisms important in learning. Activity has been conducted in the following main areas:
3.1 Measurement of Timing Requirements for Induction of Synaptic Plasticity.
Synaptic plasticity mechanisms are thought to operate at the level of individual synapses. Based on findings in other systems, the timing of presynaptic relative to postsynaptic activity is likely to be crucial in the corticostriatal system. In addition, our previous work suggests that modulation by dopamine is important. We hypothesize that the selection of appropriate corticostriatal synapses for modification during learning involves activity-dependent plasticity with precise timing requirements, which may be modulated by dopamine.
We are extending studies of dendritic mechanisms associated with spike-timing dependent plasticity (STDP) in the corticostriatal pathway. Using whole-cell recording from striatal projection neurons in adult mice we are measuring STDP over a range of different timing and excitation conditions. We have found that spike-time dependent long-term depression (tLTD) of corticostriatal inputs was maximally induced by pairing protocols in which the cortically evoked excitatory post-synaptic potential (EPSP) preceded postsynaptic action potentials by 10 ms (pre-post protocol). Post-pre protocols, or stimulation of the pre- or post-synaptic sites alone, induced no change in the cortical EPSP. the tLTD was blocked by the intracellular calcium chelators, BAPTA or EGTA, suggesting a requirement for elevated calcium for induction of tLTD. However, this form of tLTD was not blocked by the NMDA-receptor antagonist, APV, indicating a non-NMDA channel source of the calcium signal. We have now shown that calcium entry via voltage-sensitive calcium channels is necessary for tLTD (Figure 1).
Since intracellular Ca2+ levels play a key role in the induction of synaptic plasticity in the corticostriatal pathway, they may - as in other systems - be the basis of the temporal requirements observed. To investigate the mechanism underlying the spike-timing dependence of LTD, we are using 2-photon microscopy to measure intracellular Ca2+concentration in individual dendritic spines, after uncaging of glutamate adjacent to the spine. Dendritic spine Ca2+ signals have been measured during STDP induction protocols. Pre-post protocols that induced LTD were associated with larger spine Ca2+ transients than protocols that produced no LTD. These findings suggest a Ca2+ threshold for induction of LTD in the corticostriatal pathway and map out the way the timing of pre- relative to post-synaptic activity is translated into local Ca2+ signals.
Current work is investigating modulation of these timing requirements by dopamine.
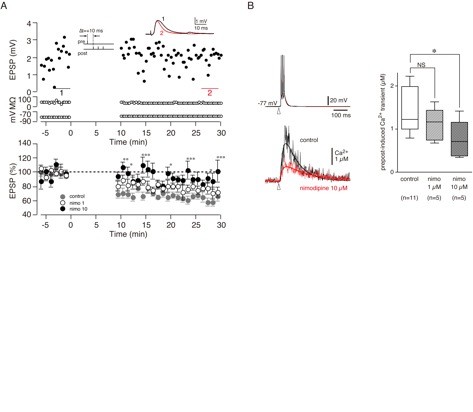
Figure 1: Supralinear [Ca2+]i increase is necessary for pre-post t-LTD and depends on voltage-sensitive calcium channels. A. Pre-post timing induced t-LTD (filled grey circles). Nimodipine 10 µM significantly blocked pre-post t-LTD (filled circles) but nimodipine 1 µM did not block pre-post t-LTD (open circles). B. Effect of nimodipine on pre-post induced Ca2+ transients. Left, upper trace shows voltage trace from recorded cell; lower trace shows comparison of Ca2+ transient under control conditions and in the presence of nimodipine 10 µM. Right, group average data shows significant effect of nimodipine 10 µM on pre-post induced Ca2+ transients.
3.2 Measurement of Cell-type Specific Differences in Synaptic Plasticity.
We have initiated studies of synaptic plasticity in striatal cells using electrical field stimulation and electrophysiological recording in brain slices from transgenic mice that express fluorescent markers in specific cell types. Transgenic mouse lines have been established, which selectively express green fluorescent protein (GFP) in subtypes of striatal spiny cells, so that we are now able to definitively identify the neurons from which records have been obtained. In the first phase of this research we developed plasticity-inducing stimulation protocols for mouse brain slices. Our ability to definitively identify morphologically similar cells by their expression of GFP is a powerful new technique that is adding important new understanding. We have tested the hypothesis that dopamine differentially regulates synaptic plasticity in dopamine D1a versus dopamine D2 receptor expressing subtypes of striatal neuron, by comparing the effects of the same induction protocols on such neurons identified by intracellular labeling and by expression of GFP. We found that STDP protocols produced LTD in dopamine D-1a receptor-expressing neurons but not in dopamine D-2 cells. However, LTD was not blocked by a D-1 receptor antagonist, indicating that although the capacity for LTD is associated with expression of the receptor, activation of the receptor is not required. This work has been completed and a manuscript is in preparation.
3.3 Optogenetic Approaches to Study Learning Mechanisms
One of the earliest issues in the theory of reinforcement learning, dating back to Thorndike, was the effectively retroactive action of reward on decision-making substrates. One idea to solve this issue was the concept of an eligibility trace, localized to synapses that were active prior to the actions that led to the reward. Subsequent reward may then act on eligible synapses, thus automatically assigning credit to neural activity that preceded the decision. Eligibility traces have been widely employed in this form in computational models of learning. We tested a synaptic eligibility trace hypothesis in corticostriatal synapses, using spike-time dependent plasticity induction protocols combined with photolytic uncaging of dopamine. In contrast to predictions, dopamine was most effective when uncaged immediately (30 ms) prior to conjunction of presynaptic and postsynaptic spikes, and had no effect when uncaged after a delay. These experiments were conducted in the dorsal striatum, and do not rule out eligibility traces in other brain regions. Also, uncaging of dopamine does not necessarily produce the same effect as the synaptic release of dopamine.
To address the actions of synaptic release of dopamine we have developed the ability to express in selected neurons the algal protein Channelrhodopsin-2, a rapidly gated light-sensitive cation channel, by using lentiviral gene delivery. We have expressed the protein in cultured striatal and cortical neurons and demonstrated light-induced activation. This technique enables temporally precise, non-invasive control of activity in well-defined neuronal populations. In ongoing work we aim to use this method to effect dopamine release by optical stimulation, which will enable us to investigate the effects of endogenously released dopamine on synaptic plasticity. To date we have been able to express channel rhodopsin 2 in dopaminergic neurons of intact mice and evoke dopamine release by optical stimulation of striatal slices prepared from the mice.
We are also using an optogenetic approach to study the involvement of specific brain regions in learning at different timepoints within each trial. In these experiments we are using halorhodopsin to inhibit neural activity in rats performing a behavioral task. By applying light at different times during each trial, we aim to dissect out the contribution of different regions to the learning task. Preliminary results show there are clear behavioral effects of inactivating the nucleus accumbens during reversal learning. We are now undertaking studies to determine the precise mechanisms of these effects.
3.4 Measurement of dopamine levels in awake animals
We have previously shown that dopamine acts at the cellular level to strengthen synapses in the striatum of the forebrain. These actions of dopamine on synaptic strength provide a cellular mechanism for reward-related learning. In this mechanism, precisely timed and pulsatile increases in dopamine concentration are critical for correct processing of reward. To complement our ongoing studies of dopamine modulation of plasticity, we aim to measure dopamine release in awake animals on a precise, subsecond timescale. We have developed the ability to use fast-scan cyclic voltammetry (FSCV) to measure dopamine release in awake-behaving rats using carbon fibre microelectrodes. We have developed hardware and software to conduct FSCV and perform principal components regression to better identify the chemicals detected. We are now using this approach to measure the phasic dopamine signal, and used this method to test a theory concerning the therapeutic actions of methylphenidate in attention-deficit hyperactivity disorder (ADHD).
Methylphenidate is the most widely used and effective treatment for ADHD but in the absence of a clear understanding of its therapeutic mechanism the use of such a powerful psychostimulant in children is controversial. We have been measuring the effects of methylphenidate on release of dopamine in an ADHD animal model and controls. We have also been studying the effects of methylphenidate on such release. These experiments have led to exciting new results that have been submitted for publication.
3.5 Theoretical and Computational Studies of the Basal Ganglia
Our electrophysiological experiments are being conducted in parallel with computational modeling of network activity in striatal inhibitory networks. We found that realistic connectivity patterns lead to spontaneous generation of assemblies that fire in sequence in response to unstructured input. This suggests an important functional role for sparse lateral inhibition in the striatum that may be relevant to neural activity sequences encoding behavior.
In addition to cellular and network modelling, we have also contributed to the development of a neurobiological hypothesis of altered reinforcement mechanisms in attention deficit hyperactivity disorder (in collaboration with the Tripp Unit). We proposed that a number of symptoms of the disorder could be interpreted as due to a failure of dopaminergic responses to established reinforcers to transfer to new predictors of reinforcement. This model makes a number of specific behavioural and neural predictions that are testable by experiment. Our published reports have been widely cited in experimental tests of the model.
4. Publications
4.1 Journals
Ponzi, A. & Wickens, J. R. Sequentially switching cell assemblies in random inhibitory networks of spiking neurons in the striatum. Journal of Neuroscience 30, 5894-5911 (2010).
Pawlack, V., Wickens, J. R., Kirkwood, A. & Kerr, J. N. Timing is not everything: neuromodulation opens the STDP gate. Frontiers in Synaptic Neuroscience 2, 1-14 (2010).
4.2 Books and other one-time publications
Wickens, J. R. & Arbuthnott, G. W. Gating of cortical input to the striatum. in Handbook of Basal Ganglia Structure and Function (eds. H. Steiner & K.Y. Tseng) (AP, 2010).
Plenz, D., Wickens, J.R. The striatal skeleton: Medium spiny projection neurons and their lateral connections. in Handbook of Basal Ganglia Structure and Function (eds. H. Steiner & K.Y. Tseng) (AP, 2010).
4.3 Oral and Poster Presentations
Wickens, J. R., Shindou, T. & Ochi-Shindou, M. Cellular mechanisms of learning in the corticostriatal system, The 5th PRESTO meeting of the research area "Decoding and Controlling Brain Information", Okinawa, Japan, Nov 5, 2010
Wickens, J. R. Cell assemblies in the neostriatal network, The Japan-Germany Workshop, Okinawa, Japan, Mar 5, 2011
Wickens, J. R. Basal Ganglia Function: Insights from Computational Modeling, IBAGS Tenth Triennial Meeting, New Jersey, USA, Jun 24, 2010
Wickens, J. R. Mechanisms of Synaptic Plasticity in the Mesostriatal System, Foundation Jean-Marie Delwart International Seminar Emotions: From neurochemistry and Brain Evolution to Human Behaviour, Genval, Belgium, Oct 16, 2010
Shindou, T., Ochi-Shindou, M. & Wickens, J. R. The dopaminergic contribution to spike-timing dependent plasticity in the corticostriatal pathway, The 87th Annual Meeting of the Physiological Society of Japan, Morioka, Japan, May 20, 2010
Shindou, T., Ochi-Shindou, M. & Wickens, J. R. Contribution of postsynaptic synapses in spike-timing dependent long-term depression of corticostriatal pathways, Neuro2010, Kobe, Japan, Sep 4, 2010
Ponzi, A. Input dependent cell assembly dynamics in an inhibitory spiking network model, Japan-Germany Joint Workshop on Computational Neuroscience, Okinawa, Japan, Mar 4, 2011
Ponzi, A. Input dependent cell assembly dynamics in a spiking model of the striatal MSN network, The 41th NIPS Sokendai International Symposium, Okazaki City, Aichi Pref, Japan, Dec 16, 2010
Ponzi, A. Input dependent cell as! sembly d ynamics in an inhibitory spiking network model, The 40th Annual Meeting of the Society for Neuroscience, San Diego, USA, Nov 14, 2010
Ponzi, A. Input dependent cell assembly dynamics in an inhibitory spiking network model, Dynamical Neuroscience XVIII, San Diego, USA, Nov 12, 2010
Ponzi, A. Input dependent cell assembly dynamics in an inhibitory spiking network model, Neuro2010, Kobe, Japan, Sep 3, 2010
Ponzi, A. Stimulus dependent population dynamics in a striatal spiking network model, Neuroinformatics 2010, Kobe, Japan, Aug 30, 2010
Ponzi, A. Input dependent population dynamics in a model of the inhibitory spiking network of medium spiny neurons in the striatum, Gaba Signalling and Brain Networks, Amsterdam, The Netherlands., Jun 30, 2010
Ponzi, A. Cell assembly dynamics in a spiking model of the striatal MSN network, OIST Junior Researcher Retreat, Okinawa, Japan, Oct 28, 2010
Ponzi, A. Input dependent cell assembly dynamics in an inhibitory spiking network model, Brain Corporation, San Diego, USA Nov 10, 2010
Ponzi, A. Investigation of Striatal Network Dynamics by Computational Modeling, IBAGS Tenth Triennial Meeting, New Jersey, USA., Jun 20, 2010
Nakano, T., Yoshimoto, J., Wickens, J. R. & Doya, K. Multi-level simulation of corticostriatal synaptic plasticity depending on input timing, The 40th Annual Meeting of the Society for Neuroscience, San Diego, USA, Nov 14, 2010
Nakano, T., Yoshimoto, J., Wickens, J. R. & Doya, K. Electrophysiological and molecular mechanisms of synaptic plasticity in the striatum, Neuro2010, Kobe, Japan, Sep 2-4, 2010
Nakano, T., Yoshimoto, J., Wickens, J. R. & Doya, K. Electrophysiological and molecular mechanisms of synaptic plasticity in the striatum: multi-scale simulation of molecule and cell., Neuroinformatics 2010, Kobe, Japan, Aug 30-Sep 1, 2010
Aquili, L. & Wickens, J. R. VTA and NAcc neurons' inhibition during reversal learning: a pharmacological and an optogenetic approach, Australian Neuroscience Society, Auckland, New Zealand., Jan 31-Feb 3 2011
5. Intellectual Property Rights and Other Specific Achievements
Nothing to report.6. Meetings and Events
6.1 OIST Seminar
Date: September 9, 2010
Venue: OIST Campus
Speaker: Dr Simon McMullan
Other remarks: Australian School of Advanced Medicine, Macquarie University, Senior Lecturer
6.2 Okinawa Computational Neuroscience Course 2010
Date: June 14-July 1, 2010
Venue: OIST Seaside House, Onna-son, Okinawa
Co-organizers: Dr Erik De Schutter, Dr Kenji Doya, Dr Klaus Stiefel, Dr Jeff Wickens, Okinawa Institute of Science and Technology
Speaker: Invited faculty
7. Others
Nothing to report.