



Information Processing Biology Unit
Principal Investigator: Ichiro Maruyama
Research Theme: Information Processing by Life

Abstract
Our unit is interested in an understanding of how cells/neurons detect external information and transmit it to the inside of the cell. For the last 20 years, ligand-induced dimerization has widely been thought to be a property common to the transmembrane signaling mechanism of all known growth factor and cytokine receptors (dimerization model). In previous years, however, we found all the members of the EGF/ErbB receptor family have preformed, yet inactive, homo- and heterodimeric structures between the family members prior to ligand binding, and have proposed an alternative 'rotation/twist' model. This fiscal year, we have determined that in living cells, the TrkA receptor for nerve growth factor (NGF) also exists as a homodimer prior to ligand binding.
We are also interested in an understanding of how the nervous system processes external information as cellular networks to regulate animal behaviors including learning and memory. Animals can survive in a narrow pH range by monitoring the pH in the environment and their body fluids. However, little is known about how animals monitor alkaline pH in the environment at molecular and cellular levels. In this study, we have employed the nematode Caenorhabditis elegans (C. elegans) as a model system. While C. elegans is attracted to low-alkaline pH, C. elegans is repelled from high-alkaline pH. We have found that GCY-14, a membrane receptor-type guanylyl cyclase, is expressed in the cilia of the ASEL amphid sensory neuron, and plays as a sensor molecule for the mild alkalinity of the environment. In this fiscal year, we have further demonstrated that a mouse homolog, mRGC, of GCY-14 is also activated by external mild-alkaline pH. In previous years, we also elucidated that OSM-9 (TRPV) channels on ASH sensory neurons play a crucial role in the sensation of strong alkalinity as a noxious stimulus. In this fiscal year, we have also found that the strong-alkalinity sensation is regulated by neuropeptides. In the past years, we also developed protocols for the study of learning and memory in C. elegans, and found that the animal can learn to associate an appetitive or aversive olfactory stimulus with an unconditioned stimulus, and can retain the associative long-term memory. This fiscal year, we have initiated to identify neural networks responsible for the learning and memory.
These results provide us insights into an understanding of a molecular mechanism underlying information transfer from the outside of cells/neurons to the inside, as well as an understanding of neural networks that control animal behaviors in response to external stimuli. These findings may also be invaluable for the development of pharmaceuticals for human diseases such as cancers and mental diseases.
1. Staff
- Hisayuki Amano, Ph.D.
- Yoko Kudeken, B.A.
- Hiraku Miyagi, M.S.
- Takashi Murayama, Ph.D.
- Hideki Nagahama, Ph.D. (until Nov. 24, 2010)
- Saori Nishijima, Ph.D.
- Shigeki Sanehisa, M.S.
- Toshihiro Sassa, Ph.D.
- Jianying Shen, Ph.D. (until Nov. 24, 2010)
- Zacharie Taoufiq, Ph.D. (until Nov. 30, 2010)
2. Collaborations
- Regulation of Cellular Functions by Signaling Molecule
- Joint research
- Prof. Ken-ichi Kariya, University of the Ryukyu School of Medicine, Okinawa, Japan
- A ubiquitin E2 variant protein acts in axon termination and synaptogenesis in Caenorhabditis elegans
- Collaborative research
- Prof. Yishi Jin, University of California, San Diego, California, USA
3. Activities and Findings
3.1. Project Aims
All forms of life are separated from non-life by cell membranes, and all cells/neurons have cell-surface receptor proteins that span the membranes in order to transmit external information, such as environmental changes and cell-cell communications, to the inside of the cell. Such information flow is fundamental for all living systems ranging from bacteria to humans. Dysregulation of the cell surface receptor molecules often causes a variety of impairments including mental and developmental diseases and cancers in humans. (1) We wish to understand at the molecular level how the external information is sensed and transmitted into the inside of cells/neurons by cell-surface receptors, and how the information is processed, transferred to other parts of the cells and regulates other cellular activities. (2) We wish also to understand information processing at higher levels through cell-cell communications; namely, how the external information is sensed and transmitted through sensory neurons, processed by the nervous system, and how it controls animal behaviors including learning and memory.
3.2. Progress report
3.2.1. Information processing by cells/neurons
3.2.1.1. Background.
The bacterial cell-surface receptor Tar recognizes aspartate molecules in the environment, and brings bacterial cells toward the higher concentration of the attractant as a nutrient, or the lower concentration of repellents such as nickel and cobalt ions. This transmembrane signaling by Tar occurs within homodimeric receptor molecules on the cell surface. We have previously shown that the Tar activity is regulated by its ligands, which bind to the extracellular domain of the receptor and lock/freeze the rotational/twist movement of the receptor’s transmembrane domains (Maruyama et al., 1995. J. Mol. Biol. 253, 530). This locking/freezing of the rotation/twist at one position by the attractant seems to inhibit the associated histidine kinase Che A, while the locking/freezing at another position by the repellent seems to activate the kinase activity (the rotation/twist model).
We have also analyzed a molecular mechanism underlying the activation of the human epidermal growth factor receptor (EGFR) family of cell-surface receptor tyrosine kinases, also known as ErbB or HER. These EGF/ErbB receptors play a pivotal role in the development of organisms, and are frequently implicated in human cancers. Furthermore, these receptors also regulate neural activities, and mutations of these receptor genes are frequently associated with mental diseases. The receptor family consists of four members, EGFR/ErbB1, ErbB2/Neu/HER2, ErbB3/HER3 and ErbB4/HER4, and has a large (~620 amino-acid residues long) extracellular ligand-binding region, a single transmembrane a-helix, and an intracellular region containing the tyrosine kinase and its regulatory domain. They form a network of homo- and heterodimers. ErbB2 can only be regulated indirectly, and is thought to be the preferred heterodimerization partner for other ErbB receptors. ErbB3, on the other hand, must associate with an ErbB family member that has an active tyrosine kinase in order to respond to its own ligand neuregulin (NRG).
Ligand-induced dimerization has widely been thought to be a property common to the transmembrane signaling mechanism of all known growth factor receptors including the EGF/ErbB receptors (Dimerization Model). According to the model, receptor dimerization is responsible for autophosphorylation of the intrinsic kinase activity, which is mediated by an intermolecular process. Namely, ligand binds to the monomeric form of the receptor, and induces its dimeric form for the activation. Prior to ligand binding, however, it still remains controversial whether the receptor has a monomeric or dimeric structure.
We have recently found by chemical cross-linking and sucrose density-gradient centrifugation that in the absence of bound ligand EGFR has an ability to form a dimer and the majority (> 80%) of the receptor exists as a preformed dimer on the cell surface. We also analyzed the receptor dimerization by inserting cysteine residues at strategic positions about the alpha-helix axis of the extracellular juxtamembrane region. The mutant receptors spontaneously formed disulfide bridges and transformed NIH3T3 cells in the absence of ligand, depending upon the positions of the cysteine residues inserted. Kinetic analysis of the disulfide bonding indicates that ligand binding induces a rotation/twist of the juxtamembrane region of the receptor in the plane parallel with the lipid bilayer. The binding of an ATP competitor to the intracellular kinase domain also induced the similar rotation/twist of the juxtamembrane region. All the disulfide-bonded dimers had flexible ligand-binding domains with the same biphasic affinities for the ligand as the wild type. Based on these results, we have proposed the ‘rotation/twist’ model for the molecular mechanism of the EGF receptor activation, in which ligand binding to the flexible extracellular domains of the receptor dimer induces the rotation/twist of the juxtamembrane regions, hence the transmembrane domains, and rearrange the kinase domains for the receptor activation. Indeed, this rotation/twist model (Figure 1; Moriki et al., 2001. J. Mol. Biol. 311: 1011; Tao and Maruyama, 2008. J. Cell Sci. 121: 3207) is consistent with the homodimeric structure of the receptor kinase, transmembrane and unactivated extracellular domains that have recently been determined by others.
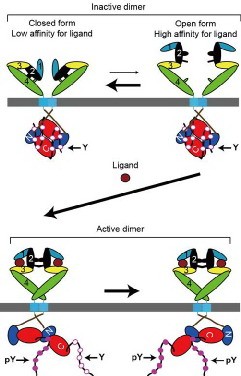
Figure 1. ‘Rotation/twist’ model for the mechanism of the ErbB receptor activation (modified from Tao & Maruyama, 2008. J. Cell Sci. 121: 3207).
To support the ‘rotation/twist’ model, we have recently determined preformed homo- and heterodimeric structures of EGFR and ErbB2 at physiological expression levels (~104 molecules per cell), using fluorescence microscopy, fluorescence resonance energy transfer (FRET) and fluorescence cross-correlation spectroscopy (FCCS) (Liu et al., 2007. Biophys. J. 93: 684). When EGFR and ErbB2 fused with a fluorescent protein (FP) were expressed on the cell surface of Chinese hamster ovary cells at physiological expression levels, FRET was detected between the donor and acceptor FPs. Furthermore, cross-correlation between FPs separately fused to EGFR or ErbB2 was also observed by FCCS, indicating that EGFR and ErbB2 molecules diffuse together as homo- or heterodimers in the cell membrane. These results demonstrate that the unactivated cell-surface receptors can spontaneously form homo- and heterodimers at physiological expression levels.
Furthermore, we have been analyzing preformed homo- and heterodimeric structures between all the members, EGFR, ErbB2, ErbB3, and ErbB4, of the receptor family by employing bimolecular fluorescence complementation (BiFC) assay, and have found that all the members display preformed, yet inactive, homo- and heterodimeric structures in the absence of bound ligand (Figure 2; Tao & Maruyama, 2008. J. Cell Sci. 121: 3207). The ligand-independent dimerization of the EGF/ErbB receptors occurs in the endoplasmic reticulum (ER) before newly synthesized receptor molecules reach the cell surface. Furthermore, we have also found that ErbB3 was localized in the nucleus when expressed alone or together with ErbB4. When coexpressed with EGFR or ErbB2, however, ErbB3 was located to the plasma membrane. These results indicate that all the EGF/ErbB receptors have homo- and heterodimeric structures before ligand binding, and are consistent with the ‘rotation/twist’ model. The ErbB receptors exist as a dimer on the cell surface, mainly through interaction between the intracellular kinase domains and C-terminal tails. The receptor dimers have flexible extracellular domains, and presumably can take two major conformations with low and high affinities for ligand. Ligand binding to the high affinity receptor stabilizes the extracellular domains, induces approximately 140-degree rotation/twist of the transmembrane domains about its helix axis in parallel to the cell membrane, dissociate the symmetric back-to-back kinase domains, and then rearrange the kinase domains to take head-to-tail asymmetric conformation for the receptor activation (Figure 1).
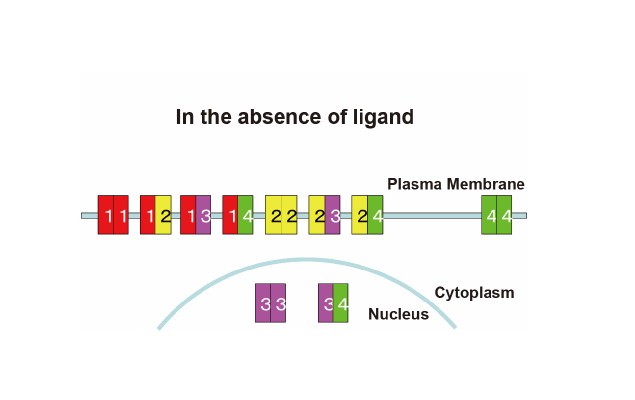
Figure 2. Preformed homo- and heterodimeric structures of the ErbB receptor family and their subcellular localization
Thus, different cell-surface receptors, bacterial Tar and human EGF/ErbB receptors, seem to be similarly regulated by their ligands in order to transmit the external information to the inside of the cell. Namely, their ligand binding regulates the rotation/twist of the receptor’s transmembrane domain in parallel to the plane of the plasma membrane. Therefore, we have been continuing to test the 'rotation/twist' model for the molecular mechanism of the activation of other cell-surface receptors including Tar, EDF/ErbB receptors and neurotrophin receptors as described below. Indeed, the neurotrophin receptor TrkA is present as a preformed, yet inactive, dimer in living cells. We have also previously found that the intracellular domain of the EGF/ErbB receptors plays a crucial role for the spontaneous formation of the receptor dimers (Tao & Maruyama, 2008. J. Cell Sci. 121: 3207). This year, therefore, we have analyzed amino-acid residues in the intracellular domain, which are involved in the preformed dimers, and have found that negatively charged amino-acid residues of the receptor carboxyl termini stabilize the preformed receptor dimers through their electrostatic interaction with the back of the kinase domains.
3.2.1.2. Crystal structure of Tar with a repellent.
The three-dimensional structures of the extracellular ligand-binding domain of the bacterial aspartate receptor Tar with or without bound ligands have been determined, and only a small (~2 Å) vertical shift of the trasmembrane domain-2 (TM2) from each other has been detected by others. This indicates that the receptor structures with and without the ligand aspartate are the same each other, consistent with the ‘rotation/twist’ model for the mechanism of the Tar activity regulation by aspartate (Maruyama et al., 1995. J. Mol. Biol. 253: 530). Furthermore, the ‘rotation/twist’ model predicts that another cofactor for Tar, nickel as a repellent in E. coli chemotaxis, stabilizes TM2 in a distinct rotational orientation against transmembrane domain-1 (TM1). Namely, the repellent nickel could induce the rotation/twist of TM2 against TM1 in parallel with the plane of the cytoplasmic membrane. To test this hypothesis, we are currently trying to determine three-dimensional structure of the extracellular domain of Tar in the presence of bound nickel.
3.2.1.3. EGFR domains required for dimerization in the absence of ligand.
Three-dimensional structures of the extracellular domains of EGFR and ErbB2 as well as of the intracellular kinase domain of EGFR recently determined by others suggest that the extracellular juxtamembrane region and intracellular kinase domain may play vital roles in the formation of the preformed dimers. Through BiFC analysis of deletion mutants, indeed, we have found that the intracellular domain of EGFR plays a crucial role for the spontaneous dimerization in the absence of ligand. Based on the three-dimensional structures, we have started systematic analysis of the EGFR domain(s) involved in the spontaneous homo- and heterodimerization. A pair of the inactive EGFR kinase domains has a symmetric back-to-back structure, in whihc its active sites face outside. The interface of the homodimer is enriched with positively charged amino acid residues, which interact with negatively charged residues of its terminal tails. We hypothesized that this ionic interaction stabilizes the homodimeric structure. By constructing point and deletion mutant molecules, therefore, we are trying to identify amino-acid residues required for the dimerization. As shown in Figure 3, this fiscal year we have found that point and deletion mutations of the negatively charged residues of the EGFR regulatory tail spontaneously activate the receptor. These results are consistent with our hypothesis that the ionic interaction at the dimer interface stabilizes the dimeric structure. These results support the 'rotation/twist model', not the 'dimerization model'.
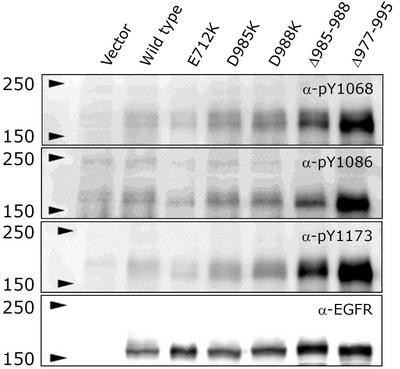
Figure 3. Ligand-independent activation of EGFR mutants with a point or deletion mutation. Amino acid residues residing in the interface of the symmetric dimeric structure were replaced or deleted. Note that the mutant receptors were activated in the absence of bound EGF.
3.2.1.4. Preformed, yet inactive, dimeric structures of receptors for neurotrophic factors.
The nerve growth factor (NGF) family of neurotrophins that includes NGF, brain-derived neurotrophic factor (BDNF), neurotrophin-3 (NT-3), and NT-4/5 play a central role in the development, growth, and maintenance of the nervous system. These neurotrophins interact with two types of cell surface receptors, the tropomyosin-related kinase (Trk) family of tyrosine kinase receptors, TrkA, TrkB and TrkC, and the p75 neurotrophin receptor (p75NTR). Trk receptor subtypes bind mature neurotrophins with different specificities: TrkA and TrkC preferentially bind NGF and NT-3, respectively, while TrkB mainly binds BDNF and NT-4. The p75NTR binds all mature neurotrophins with approximately equal low affinity and has been demonstrated to bind the proneurotrophins with high affinity. These receptors are present on the same cell, and modulate the responses of neurons to neurotrophins.
NGF-induced activation of TrkA is thought to be mediated by receptor dimerization (dimerization model). Cross-linking studies revealed the formation of TrkA homodimers upon NGF binding to pheochromocytoma PC12 cells and fibroblasts ectopically expressing TrkA receptors. Because NGF, as well as other neurotrophins, exist in solution as stable homodimers, it is thought that a single NGF dimer bridges two TrkA monomers. This model was supported by the symmetric crystal structure of the NGF complex with the extracellular IgG domain of TrkA. However, NGF mimetics and Fab fragments of antibody against TrkA, which are structurally incapable of dimerizing the receptor, could activate TrkA. Furthermore, an asymmetric chimeric NGF/Neurotrophin-4 heterodimer also was found to activate TrkA receptor. TrkA receptor molecules expressed in Xenopus Laevis oocytes were found to be present as oligomers in the plasma membrane in the absence of NGF. These previous results led us to examine whether neurotrophin receptors exist as monomers or preformed dimers on the cell surface prior to ligand binding.
In the past years and this fiscal year, we have analyzed structures of the TrkA receptor for NGF by chemical crosslinking, bimolecular fluorescence complementation (BiFC) and luciferase fragment complementation assays. These analyses demonstrated that before ligand binding, TrkA exists as homodimers in living cells (Shen & Maruyama, FEBS Lett. 585, 295-299, 2011). Using Brefeldin A, which disassembles the Golgi apparatus and blocks anterograde transport of the receptors from endoplasmic reticulum (ER) to Golgi, furthermore, it was found the preformed dimers were formed in ER before reaching Golgi. This work provides new insights into understanding of transmembrane signaling by receptors for neurotrophic factors.
3.2.2. Information processing by the nervous system
3.2.2.1. Alkalinity sensation in C. elegans.
3.2.2.1.1. Background.
Like other animals, the nematode Caenorhabditis elegans detects various environmental cues such as tastes and odors mainly through its amphid sensilla. The amphids are the largest chemosensory organs, and each amphid includes 12 sensory neurons (ADF, ADL, AFD, ASE, ASG, ASH, ASI, ASJ, ASK, AWA, AWB, and AWC) with ciliated dendrites as well as one sheath and one socket cell. Apart from AFD, the sensory dendrites of 11 neurons penetrate the sheath-cell ending, and the cilia of eight of these neurons, except those of AWA, AWB, and AWC, extend into the doughnut-like pore created by the socket cell and are directly exposed to the external medium. These amphid neurons have roles in chemotaxis, thermotaxis, mechanosensation, osmotaxis, and dauer pheromone sensation.
Animals can survive in a narrow pH range, and continuously monitor the pH of environment and body fluids. However, little is known about how animals monitor alkaline pH in the environment, although channels that can sense intracellular alkaline pH have recently been found. Environmental mild alkalinity is one of attractive cues for C. elegans. Between pH 3.0 to pH 10.0, wild-type animal prefers higher alkaline pH ranges, while the animal avoids higher alkalinity than pH 11.0 and acidic conditions lower than pH 4.0. However, neurons and cell-surface molecules that sense the low and high alkaline pH ranges have not been elucidated in C. elegans. Neural networks responsible for worm’s attraction to mild alkalinity and aversion from strong alkalinity can also be efficiently analyzed since the wiring diagrams of all the neurons have been reconstructed from electron micrographs of serial thin sections of the C. elegans whole body.
3.2.2.1.2. Mild-alkalinity sensation.
In previous years, we have devised an agar plate assay with a linear pH gradient in order to investigate cellular and molecular bases for C. elegans chemotaxis toward low-alkaline pH. Along the pH gradient from pH 6.8 to pH 8.5, wild-type animals were attracted to higher pH regions, whereas che-1 mutants defective in chemosensory ASE neurons were not. To search for a molecular sensor for the low-alkaline pH, we have performed a series of RNAi knock down of genes encoding various channels, cell-surface receptors and guanylyl cyclases in conjunction with analysis of chemotaxis mutants previously known. Among the genes analyzed, tax-2, tax-4 and gcy-14 have been found to be involved in the C. elegans chemoattraction to the low-alkaline pH. By imaging of Ca2+ concentration changes using a Ca2+-sensing fluorescent protein, furthermore, we found that ASE-left (ASEL) is activated by pH up-step. GFP-tagged GCY-14 was localized to ASEL sensory cilia, and in the gcy-14 mutant, ASEL did not respond to pH up-step. While ASI and ASK sensory neurons did not respond to pH up-step, furthermore, ASI and ASK neurons ectopically expressing GCY-14 were activated by pH up-step from pH 7.0 to pH 10.0. This demonstrates that GCY-14 is sufficient for the alkaline pH sensation. These results indicate that the transmembrane guanylyl cyclase GCY-14 acts as an alkaline pH sensor, and an increased concentration of cGMP opens the cGMP-gated cation channel TAX-2/TAX-4 for the activation of ASEL sensory neurons.
This fiscal year, we analyzed receptor-type guanylyl cyclase (RGC) homologs of the C. elegans GCY-14 in mice. When Chinese hamster ovary (CHO) cells expressing mRGC was incubated with alkaline pH buffer, an intracellular cGMP level was increased as the external pH values were raised up to pH 9.0 (Figure 4A). This result suggests that mRGC might be an ortholog of GCY-14 in mice, which play as an alkalinity sensor for external alkaline pH. When the cells were incubated with mild-acidic buffer containing NH4Cl, which induced intracellular pH to mild-alkaline pH, by contrast, no increase in an intracellular concentration of cGMP was observed (Figure 4A). These results demonstrate that mRGC detects extracellular, not intracellular, alkaline pH, and results in producing cGMP inside the cell.
When mRGC was expressed in the C. elegans ASEL chemosensory neuron lacking GCY-14, furthermore, the ASEL was activated by environmental alkaline pH. This demonstrate that mRGC can play a role as an extracellular alkaline pH sensor by replacing GCY-14 (Figure 4B).
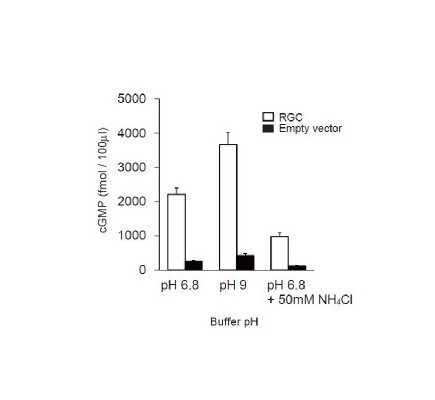
Figure 4. Mouse RGC functions as an extracellular alkalinity sensor. cGMP levels in CHO cells expressing mRGC (open bar), or empty vector (filled bar), respectively.
3.2.2.1.3. Strong-alkalinity sensation.
During the work on mild-alkalinity sensation described above, we have found that C. elegans avoids higher pH than 11.0 using a quadrant-plate assay (Figure 5A). In previous years, we have identified several genes that play pivotal roles in the avoidance behaviour: che-2, dyf-3, osm-1 and osm-5 required for chemosensation, and osm-9 and ocr-2 encoding subunits of the transient-receptor potential type-V (TRPV) channel. Rescue experiments of dyf-3 and osm-9 mutants by injecting the respective cDNA into the chemosensory neuron ASH indicate that ASH is a major neuron responsible for the high alkaline pH sensation in C. elegans. This was confirmed by Ca2+ imaging in which upon stimulation with high alkaline pH, a Ca2+ concentration increase was observed in ASH (Figure 5C)
In this fiscal year, we have also found that goa-1, which encodes an alpha–subunit of heterotrimeric G-proteins as well as egl-3 and egl-21 encoding enzymes responsible for vesiclular neuropeptide processing play a pivotal role in the avoidance behaviors. Increases of Ca2+ concentrations in ASH neurons of goa-1, egl-3 and egl-21 mutants were normal upon high alkaline pH stimulation (Figure 5D), indicating that Ca2+ influx into ASH is not affected by these mutations.
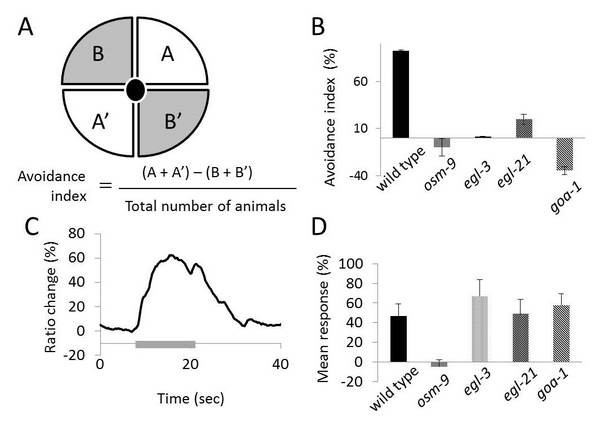
Figure 5. (A) Quadrant-plate assay. A and A’ regions are at pH 7.0, and B and B’ at pH 11.0. (B) Mutants defective in avoidance response from high alkaline pH. (C) Ca2+ imaging of ASH upon stimulation with high alkaline pH (grey bar). (D) Mean responses of ASH neurons to environmental high-alkaline pH in various mutants indicated, which were analyzed by Ca2+ imaging.
3.2.2.2. Associative learning and memory in C. elegans
3.2.2.2.1. Background.
C. elegans is an excellent model organism for the study of learning and memory. The hermaphrodite nervous system consists of only 302 neurons, and the neural circuits of these invariant neurons have completely been reconstructed from electron micrographs of serial thin sections of the entire body. The worm body is transparent throughout its life so that neural activities can directly be observed using Ca2+-sensitive fluorescent proteins in living animals.
Associative learning in C. elegans has first been suggested from the finding that worms return to their temperature of cultivation if they had food at that temperature. Most of learning paradigms in C. elegans are based on pairing chemical cues or cultivation temperature with food or starvation. Rather than pairing chemical cues with food or starvation, however, it would be preferable for subsequent analysis of neural circuits responsible for associative learning and memory that two defined chemical cues are used for conditioning of animals in associative learning paradigms.
3.2.2.2.2. Appetitive olfactory learning and associative long-term memory
In previous years, we have developed protocols for classical (Pavlovian) conditioning of C. elegans with nonanol, as a conditioned stimulus (CS), and potassium chloride (KCl) as an unconditioned stimulus (US). Before the conditioning, animals avoided nonanol, an aversive olfactory stimulus, and were attracted by KCl, an appetitive gustatory stimulus, in chemotaxis assay. After eight-cycle massed (without intertrial intervals, ITI) or spaced (with 10-min ITI) training, in contrast, animals were attracted to nonanol. Memory induced by the massed training was no longer observable within three hours, while the spaced training induced the memory that was retained for more than 12 hours. Animals treated with cycloheximide or actinomycin D failed to form the long-lasting memory by the spaced training, whereas the memory induced by the massed training was not significantly affected. These results indicated that the memory formation by the spaced training, but not by the massed training, required protein synthesis and mRNA transcription. Therefore, the memories induced by the massed and spaced training are classified as short-term/middle-term (STM/MTM) and long-term (LTM) memories, respectively. In support of this, C. elegans mutants defective in nmr-1 encoding an NMDA receptor subunit failed to form both of the STM/MTM and LTM, while mutations in crh-1 encoding the CREB transcription factor affected only the formation of the LTM.
This fiscal year, we have tried to determine sensory neurons that detect environmental nonanol in C. elegans toward the elucidation of neural networks responsible for the associative learning and memory between nonanol and KCl. Olfactory avoidance assays of animals whose amphid AWB or ASH neurons were separately killed by laser microsurgery demonstrate that AWB is responsible for the perception of a low concentration, 0.1%, of nonanol (Figure 6).
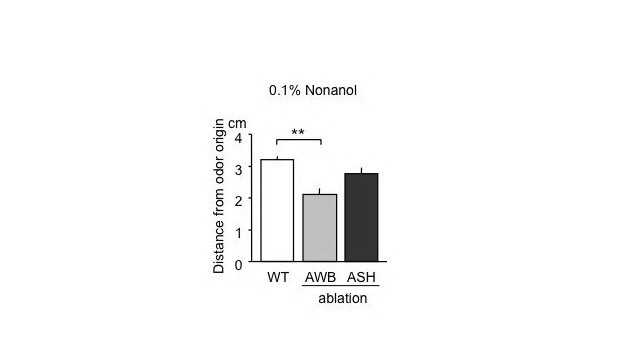
Figure 6. Nonanol avoidance of animals with laser microsurgery of sensory neurons. Animals were placed at the center of agar plates, 6 cm in diameter, where nonanol solution was spotted 1.5 cm away from the center. Distances of animals from the nonanol origin were measured in10 min. Note that animals with laser ablation of AWB were less sensitive to 0.1% nonanol that that of wild-type animals.
3.2.2.2.3. Aversive olfactory learning and associative long-term memory
In previous years, we have continuously been developing protocols for classical conditioning of animals with propanol, as CS, and hydrochloric acid (HCl), pH 4.0, as US. Before the conditioning, worms were attracted to propanol, and avoided HCl in chemotaxis assay. After spaced or massed training, by contrast, animals were either not attracted at all or were repelled from propanol on the assay plate. The memory after the spaced training was retained for 24 h, while the memory after the massed training was no longer observable within 3 h. Animals pretreated with transcription and translation inhibitors failed to form the memory by the spaced training, whereas the memory after the massed training was not significantly affected by the inhibitors and was sensitive to cold-shock anesthesia. Furthermore, the memory after spaced training was partially disrupted by extinction learning, which is repeated exposed to the CS in the absence of US. Therefore, the memories after the spaced and massed trainings can be classified as LTM and STM/MTM, respectively. Consistently, like other organisms including Aplysia, Drosophila, and mice, C. elegans mutants defective in nmr-1 encoding an NMDA receptor subunit failed to form both LTM and STM/MTM, while mutations in crh-1 encoding the CREB transcription factor affected only the LTM (Amano & Maruyama, 2011. Learn. Mem. in press).
4. Publications
4.1 Journals
Miyagi, H., Maruyama, I. N. Analysis of ligand-receptor interaction on the surface of living cells by fluorescence correlation spectroscopy. The Open Spectroscopy Journal 4, 28-31 (2010).
Trujillo, G., Nakata, K., Yan, D., Maruyama, I. N., Jin, Y. A ubiquitin E2 variant protein acts in axon termination and synaptogenesis in Caenorhabditis elegans. Genetics 186, 135-145 (2010).
Shen, J., Maruyama, I. N. Nerve growth factor receptor TrkA exists as a preformed, yet inactive, dimer in living cells. FEBS Lett 585, 295-299 (2011).
Amano, H., Maruyama, I. N. Aversive olfactory learning and associative long-term memory in Caenorhabditis elegans. Learning & Memory (2011) in press.
4.2 Oral and Poster Presentations
Shen, J., Maruyama, I. The nerve growth factor receptor TrkA exists as a pre-formed homodimer before ligand binding, Experimental Biology 2010 (International), Anaheim Convention Center, Anaheim, California, USA, Apr 27, 2010.
Maruyama, I. ‘Rotation/twist’ model for the molecular mechanism of the EGF/ErbB receptor family activation, 3rd World Cancer Congress 2010 (International), Expo Convention and Exhibition Center, Singapore, Jun 22, 2010.
Murayama, T., Fujiwara, M., Takayama, J., Maruyama, I. Alkaline pH sensation mediated by a Caenorhabditis elegans transmembrane guanylyl cyclase, C. elegans Topic Meeting 2010 'Neuronal Dvelopment, Synaptic Functin & Behavior' (International), University of Wisconsin-Madison, Memorial Union, 800 Langdon Street, Madison, Wisconsin 53706, USA, Jun 27, 2010.
Amano, H., Maruyama, I. Associative learning and memory in Caenorhabditis elegans, C. elegans Topic Meeting 2010 'Neuronal Dvelopment, Synaptic Functin & Behavior' (International), University of Wisconsin-Madison, Memorial Union, 800 Langdon Street, Madison, Wisconsin 53706, USA, Jun 28, 2010.
Amano, H., Maruyama, I. Associative learning and memory retention in Caenorhabditis elegans, 4th East Asia C. elegans Meeting (International), National Olympic Memorial Youth Center, Tokyo, Japan, Jul 12, 2010.
Nishijima, S., Amano, H., Maruyama, I. Reward learning and long-term memory in C. elegans, 4th East Asia C. elegans Meeting (International), National Olympic Memorial Youth Center, Tokyo, Japan, Jul 12, 2010.
Murayama, T., Fujiwara, M., Takayama, J., Maruyama, I. Alkaline pH sensation mediated by a Caenorhabditis elegans transmembrane guanylyl cyclase, 4th East Asia C. elegans Meeting (International), National Olympic Memorial Youth Center, Tokyo, Japan, Jul 13, 2010.
Sassa, T., Sanehisa, S., Murayama, T., Maruyama, I. Molecular and cellular bases for high-alkaline pH avoidance in C. elegans, 4th East Asia C. elegans Meeting (International), National Olympic Memorial Youth Center, Tokyo, Japan, Jul 13, 2010.
Amano, H., Maruyama, I. Associative learning and memory retention in Caenorhabditis elegans, Neuro 2010 (Domestic in English), Kobe Convention Center, Kobe, Hyogo, Japan, Sep 2, 2010.
Nishijima, S., Amano, H., Maruyama, I. Long-term appetitive memory in the nematode C. elegans, Neuro 2010 (Domestic in English), Kobe Convention Center, Kobe, Hyogo, Japan, Sep 2, 2010.
Sassa, T., Sanehisa, S., Murayama, T., Maruyama, I. Molecular and cellular bases for high-alkaline pH avoidance in C. elegans, Neuro 2010 (Domestic in English), Kobe Convention Center, Kobe, Hyogo, Japan, Sep 2, 2010.
Murayama, T., Fujiwara, M., Takayama, J., Maruyama, I. Alkaline pH sensation mediated by a Caenorhabditis elegans transmembrane guanylyl cyclase, Neuro 2010 (Domestic in English), Kobe Convention Center, Kobe, Hyogo, Japan, Sep 4, 2010.
Nishijima, S., Maruyama, I. Appetitive olfactory learning and long-term associative memory in C. elegans, Neuroscience 2010 (International), San Diego Convention Center, San Diego, CA, USA, Nov 15, 2010.