



G0 Cell Unit
Principal Investigator: Mitsuhiro Yanagida
Research Theme: Cellular Strategy for Maintaining Starved G0 Arrest and Promoting Vegetative Proliferation

Abstract
The regulation of cellular proliferation and quiescence (G0 phase) in response to changes in the environment, such as the availability of nutrients, is a central issue in biology and medicine. Our research goal is to understand the molecular mechanisms by which cell division and cell cycle arrest in the G0 phase are regulated in response to available nutrients. To this end, we have adopted the fission yeast Schizosaccharomyces pombe, a simple unicellular eukaryote, as a model organism. This year, we continued to study G0 phase induced by nitrogen starvation and adaptation of S.pombe to glucose-limited condition. First, the comprehensive mutant screen was performed to identify genes, which are non-essential for cell division in the complete medium but become essential for quiescence under nitrogen starvation. Second, we analyzed previously identified genes required for G0 phase maintenance. The ubiquitin/proteasome system and autophagy cooperatively maintain mitochondria and prevent the lethal accumulation of reactive oxygen species (ROS). Finally, we investigated cell division-quiescence behavior of the fission yeast S. pombe under a wide range of glucose concentrations. The metabolomic changes under glucose-limited condition were revealed by the mass spectrometric method established in 2009. Furthermore, in collaboration with Kyoto University, regulations on mitotic chromosomes were investigated in 2010. An unexpected link between nutritional control and kinetochore function, and essential roles of Condensine throughout mitosis were reported. Comprehensive screening was continued to identify genes affecting adaptation to low glucose condition. Based on these and previopus studies, we published five original research articles and two reviews in 2010.
1. Staff
- Dr. Kojiro Takeda, Group leader
- Dr. Takahiro Nakamura, Researcher
- Dr. Kumiko Ohta, Researcher
- Dr. Kenichi Sajiki, Researcher
- Dr. Takeshi Hayashi, Researcher (Kyoto)
- Dr. Norihiko Nakazawa, Researcher (Kyoto)
- Ms. Sakura Kikuchi, Technical staff
- Ms. Risa Uehara, Technical staff
- Ms. Ayaka Mori, Technical staff
- Mr. Tomas Pluskal, Technical staff
- Mr. Alejandro Villar-Briones, Technical staff
- Ms. Tomomi Teruya, Research administrator/Secretary
- Ms. Takako Shiono, Research administrator/Secretary (Kyoto)
- Ms. Yuria Tahara, Research assistant
- Ms. Eulaia, Lopez-Galcia, Research assistant
2. Collaborations
- Theme: Metabolite structure determination using NMR
- Type of collaboration: Joint research
- Researchers:
- Professor Masaru Ueno, Department of Molecular Biotechnology, Graduate School of Advanced Science of Matter, Hiroshima University
- Theme: Identification of fission yeast mutants, which show growth defect under low glucose condition
- Type of collaboration: Joint research
- Researchers:
- Professor Shigeaki Saitoh, Institute of Life Science, Kurume University
- Theme: Metabolomic analysis of human cultured cells under low-glucose condition
- Type of collaboration: Joint research
- Researchers:
- Professor Hideo Orimo, Division of Metabolism and Nutrition, Department of Biochemistry and Molecular Biology, Nippon Medical School
- Dr. Asako Kounosu, Division of Metabolism and Nutrition, Department of Biochemistry and Molecular Biology, Nippon Medical School
- Theme: Identification of the molecular mechanism, required for the maintenance of cell viabiity in low glucose condition
- Type of collaboration: Joint research
- Researchers:
- Professor Kunihiro Ohta, Department of Life Science, Graduate School of Arts and Science, University of Tokyo
- Dr. Tomoichiro Miyoshi, Department of Life Science, Graduate School of Arts and Science, University of Tokyo
- Theme: Metabolomic analysis of human blood
- Type of collaboration: Joint research
- Researchers:
- Dr. Hiroshi Kondoh, Department of Geriatric Medicine, Graduate School of Medicine, Kyoto University
- Dr. Takumi Mikawa, Department of Geriatric Medicine, Graduate School of Medicine, Kyoto University
- Theme: Understanding of the molecular mechanism for regulating cell quiescence and for responding changes of nutrient condition
- Type of collaboration: Joint research
- Researchers:
- Dr. Nobuyasu Ikai, Laboratory of Molecular Inheritance, Graduate School of Biostudies, Kyoto University
3. Activities and Findings
3.1 Identification of Genes Essential for G0 Entry and/or Maintenance under Nitrogen Starvation
Transition from proliferation to quiescence brings about extensive changes in cellular behavior and structure. However, essential genes for quiescence establishment and maintenance are largely unknown. The fission yeast S. pombe is an excellent model for studying this problem, because it becomes quiescent under nitrogen starvation. Also in recent years, deletion mutant library sets have become commercially available (Bioneer Corp. Korea). We used the Bioneer haploid deletion set to identify genes that are non-essential for cell division in the complete medium but become essential for quiescence under nitrogen starvation. By spot testing and viability screening under nitrogen starvation, we analyzed more than 2800 deletion strains, using two laboratory automation systems to handle with the number of strains (Figure 1).
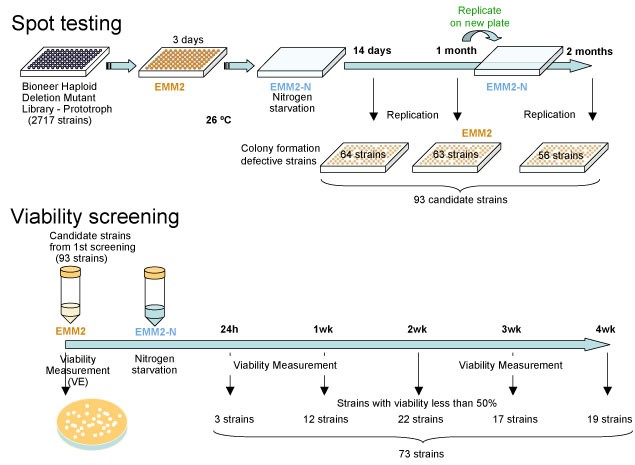
Figure 1: The strategy of screening mutants that have defects in G0 maintenance.
From these screenings, we identified 73 strains that considerably dropped the viability within 4 weeks after nitrogen starvation. The deleted genes of these strains covered cellular functions of autophagy, endosome-vacuole trafficking, exocytosis, mitochondria, phosphate signaling, chromatin remodeling, transcription factors and so on. Among them, protein phosphatase regulators are significantly abundant. Their absence caused the abnormal morphology after nitrogen starvation like septated and chitin accumulated cells and large vacuole containing cells with no polarity.3.2 Essential Roles of the Proteasome in G0 Phase ans Proliferation
While the ubiquitin/proteasome system is pivotal to cell regulations, its essential roles in the G0 phase maintenance are elusive. As the proteasome is a target of chemical anti-cancer therapy (anti-cancer drug Velcade/Bortezomib is a specific inhibitor for the ptoteasome), it is important to deepen our knowledge on the biology of the proteasome pathway in both proliferatio and the G0 phase. Our recent study suggests that the proteasome may, in cooperation with autophagy, contribute to life span the G0 phase through quality control of mitochondria, the cellular powerhouse (Figure 2). The proteasome is important to maintain mitochondria and autophagy system scavenges damaged mitochondria, which are accelerated in ROS (reactive oxygen species) production. These results were published in Takeda et al (PNAS, 2010, original research article) and in Takeda and Yanagida (Autophagy, 2010, review article).
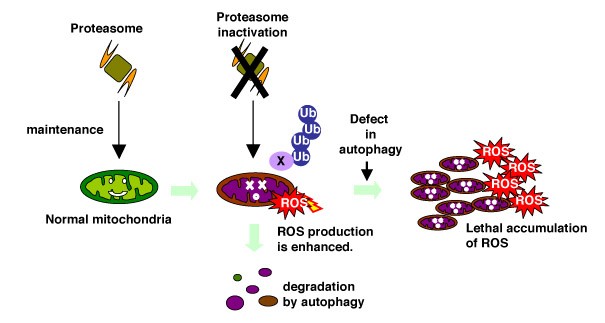
Figure 2: The model of cooperation of the proteasome and autophagy in G0 phase.
The ubiquitin/proteasome system may be engaged in maintenance of mitochondria and autophagy scavenges damaged mitochondria in order to reduce harmful ROS accumulation.
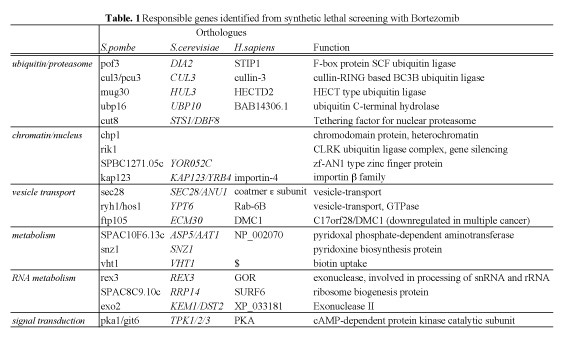
To further our knowledge in the proteasome biology in proliferation, we aimed to identify S.pombe genes affecting the cytotoxity of anti-cancer drug Bortezomib by genome-wide synthetic lethal screening. Among the 2815 genes screened, 19 genes, whose deletions induce strong synthetic lethality with Bortezomib, were identified. We designated such genes as SLB (Synthetic Lethal with Bortezomib) genes (Table. 1). Thirteen identified genes are conserved in human and thus potentially interesting for further studies in human culture cells. If the same synthetic lethality effects occur in human cells, such SLB genes have potential for the innovation of new therapies or diagnosis. These results will be published in 2011.
3.3 S.pombe Metabolomic Analysis under Low Glucose
In the fiscal year 2010, we investigated cell division-quiescence behavior of the fission yeast S. pombe under a wide range of glucose concentrations (0-111 mM; Figure 3A). The study was published in FEBS Journal (Pluskal et al., 2011). The S. pombe cells were fixed in a microfluidic perfusion system, which was constantly supplied with fresh culture medium. The mode of cell division was surprisingly normal under highly diluted glucose concentrations (5.6 mM, 1/20 of the standard medium, within the human blood sugar levels). Division became stochastic in 2.2-4.4 mM glucose. A critical transition from division to quiescence occurred within a narrow range of concentrations (2.2-1.7 mM). Under starvation (1.1 mM), cells were mostly quiescent and only a small population of cells divided. Under fasting (0 mM) condition, division was immediately arrested with a short chronological lifespan (Figure 3B). However, when cells were first glucose starved prior to fasting, they possessed a substantially extended lifespan (~14 days).
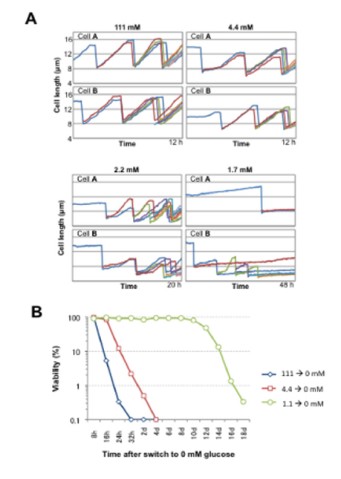
Figure 3: A. Growth of S.pombe cells cultivated in EMM2 media with reduced glucose content. Cell length increase over time is shown. B. Viability of S.pombe cells cultivated in a medium completeklu lacking glucose, depending on the initial gkucose concentration before the medium switch.
We employed quantitative metabolomic approach for cell extracts using LC-MS, and identified specific metabolites (e.g., biotin, trehalose, ergothioneine, S-adenosyl methionine, and CDP-choline), which increased or decreased at different glucose concentrations, while nucleotide triphosphates such as ATP kept high concentrations even under starvation (Figure 4).
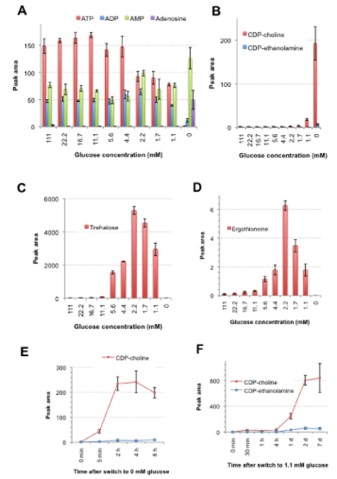
Figure 4: Peak areas of potential biomarker compounds in 10 different concentrations determined by the LC/MS method. Metabolite extracts were prepared three times and mean peak areas with standard deviations of the following metabolites are shown: A. ATP, ADP, AMP and adenosine. B. CDP-choline, CDP-ethanolamine. C. trehalose. D. ergothioneine. E. The time course change of the peak areas of CDP-choline and CDP-ethanolamine in cells switched to a fasting (0 mM) glucose condition from 111 mM glucose. F. The time course change of the peak areas of CDP-choline and CDP-ethanolamine in cells switched to the starvation condition (1.1 mM glucose) from 111 mM glucose.
Considering the rapid decrease of the antioxidants glutathione and ergothioneine (a mitochondrial antioxidant) in low glucose, we employed a fluorescent probe H2DCFDA to check for the presence of oxidative stress. Only cells abruptly shifted from 111 mM to 0 mM glucose for 6 h had strong fluorescent signals (Fig. 5A). We challenged the cells with 40 mM H2O2, a concentration previously reported to kill S. pombe cells within one hour. Results (Fig. 5B) indicate that cells incubated in 0 mM glucose for 6 h (blue rectangles) were sensitive to H2O2 oxidative stress, while cells cultivated in 1.1 mM glucose (green rectangles) were much more resistant than cells in 111 mM glucose (red rectangles) or 0 mM fasting cells. Fasting cells abruptly shifted from 111 mM glucose were the most sensitive to 40 mM H2O2. Based on the cellular and metabolic phenotypes obtained, we designated the different glucose concentrations excess (111 mM), regular (11.1 mM), diet (4.4 mM), severe diet (2.2 mM), sub-starvation (1.7 mM), starvation (1.1 mM), and fasting (0 mM), as summarized in Fig. 5C.
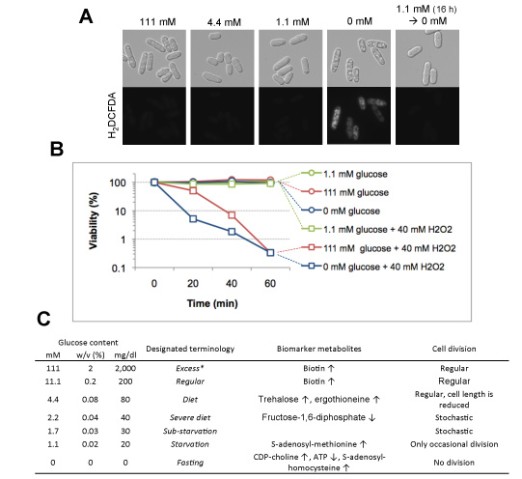
Figure 5. A. Oxidative stress accumulation was observed using the H2DCFDA dye. B. Cells were incubated in excess (111 mM), starvation (1.1 mM) and fasting (0 mM) glucose conditions for 6 h, followed by the addition of H2O2 to the final concentration of 40 mM. }Viability (%) was measured in 20 min intervals. C. Summary of the glucose concentrations described in this manuscript. Concentrations are listed in three commonly used notations: millimolar (mM), percentage (w/v), or mg/dl. Biomarker metabolites increased (↑) or decreased (↓) in each condition are noted, as well as the corresponding cell division phenotypes.
3.4 A Possible Relationship between Kinetochore Functions and Nutritional Control
In collaboration with Kyoto University, an unexpected link between nutritional control and centromere/kinetochore functions was discovered and reported in PLoS One journal (Shiroiwa et al., 2011). The centromere is the chromosome domain on which the mitotic kinetochore forms for proper segregation. Deposition of the centromeric histone H3 (CenH3, CENP-A) is vital for the formation of centromere-specific chromatin. The Mis6-Mal2-Sim4 complex of the fission yeast S. pombe is required for the recruitment of CenH3 (Cnp1), but its function remains obscure. We performed mass spectrometric analysis of the Mis6 and Mis17 complex. The results together with the previously identified Sim4- and Mal2-TAP precipitated proteins indicated that the complex contains 12 subunits, Mis6, Sim4, Mal2, Mis15, Mis17, Cnl2, Fta1-4, Fta6-7, nine of which have human centromeric protein (CENP) counterparts. Furthermore, we found that Mis17 was highly phosphorylated (Figure 6). Domain dissection of Mis17 indicated that the carboxy-half of Mis17 is functional, while its amino-half is regulatory. Overproduction of the amino-half caused strong negative dominance, which led to massive chromosome missegregation and hypersensitivity to the histone deacetylase inhibitor TSA. Negative dominance of the Mis17 fragment is exerted while the complex and CenH3 remain at the centromere. This finding is distinct from the mislocalization or loss phenotype found in ts mis17-362 cells. Overproduction-induced negative dominance was abolished in actin- and nutrition-related kinase-deletion mutants, ssp2 (AMPK), ppk9 (AMPK), ppk15 (Yak1), ppk30 (Ark1), wis4 (Ssk2), and lsk1 (P-TEFb) (Figure 6). These findings suggest an unexpected link between Mis17 and control of the cortex actin, nutrition, and signal/transcription (Figure 6).
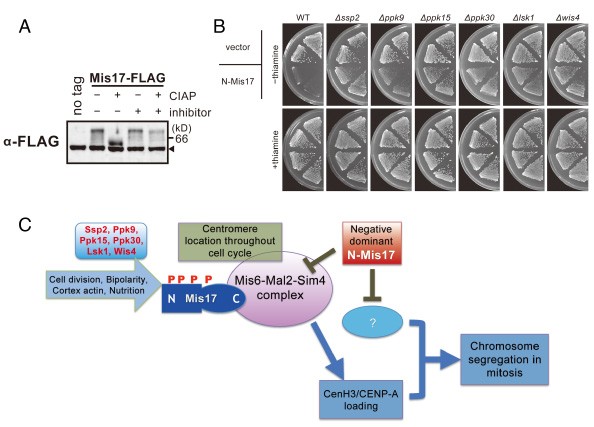
Figure 6: Mis17 is a regulatory module of the Mis6 complex
A. Mis17 is a hyperphosphoprotein. B. The negative dominance effect of N-Mis17 overexpression was greatly diminished in six protein kinase deletion mutants, ∆ssp2, ∆ppk9, ∆ppk15, ∆ppk30, ∆lsk1, and ∆wis4. C. A cartoon for the implication of Mis17 in the Mis6-Mal2-Sim4 complex and negative dominance effect of the N-Mis17 fragment.
3.5 Roles of Condensin throughout Mitosis
Condensin is a conserved protein complex that functions in chromosome condensation and segregation. It has not been unequivocally determined whether condensin is required throughout mitosis. We critically examined whether S.pombe condensin continuously acts on chromosomes during mitosis and compared the role with that of DNA topoisomerase II (Top2). Temperature shift-up experiments were done using double mutants containing the temperature-sensitive cut14-208 (SMC2) or top2 mutations combined with the cold-sensitive mutant nda3-KM311 (b-tubulin). These experiments allowed inactivation of condensin or Top2 even after late anaphase. The results established that mitotic chromosomes require condensin and Top2 throughout mitosis, and even in telophase (Figure 7A). We then show that Cnd2/Barren is the target subunit of aurora-B like kinase Ark1. Phosphorylation sites in Cnd2 were determined by mass spectrometry and their alanine and glutamate substitutions were constructed. The Ark1-mediated phosphorylation of Cnd2 S52 occurred throughout mitosis (Figure 7B). Alanine mutants of Cnd2 exhibited broad mitotic defects, including telophase, and also severe dominant-negative effect when overproduced. By contrast, glutamate mutants alleviated segregation defect in Ark1-inhibited cells. Mutant condensin in telophase accumulated in ‘lump’ that contained telomeric DNA and proteins that failed to segregate. Condensin may thus have to keep apart the segregated chromosomes in telophase. This study was published in the Journal of Cell Science (Nakazawa et al., 2011).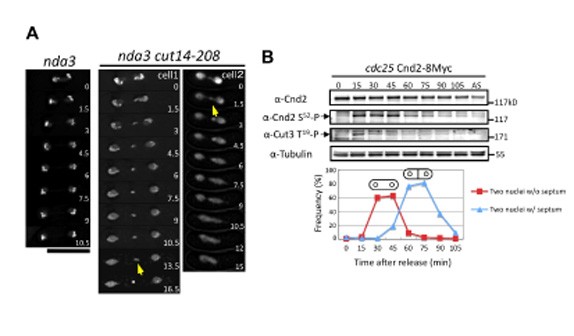
Figure 7: Continuous requirement of condensin throughout mitosis
A. Inactivation of condensin after entry into anaphase causes defects in telophase. The arrows indicate the lump structure. The numbers in the panels indicate time in minutes. Bar, 10 mm. B. Cnd2 S52 residue is phosphorylated throughout mitosis. The cdc25-22 Cnd2-8Myc cells were arrested at late G2 at 36°C and then released at 26°C. In contrast to the Cnd2 S52, the phosphorylation of Cut3 T19 briefly peaked only at 15 minutes, corresponding to prophase and metaphase.
3.6 Other Activities in Progress
Comprehensive screening mutants showing growth defect under glucose-limited condition was performed and functional analyses on isolated mutants were in progress (in collaboration with Kurume University).
4. Publications
4.1 Journals
- Yanagida, M. Mostly DNA, a bit of glucose, and the next 50 years. Mol Biol Cell 21, 3826, doi:21/22/3826 [pii] 10.1091/mbc.E10-04-0367 (2010).
- Takeda, K. & Yanagida, M. In quiescence of fission yeast, autophagy and the proteasome collaborate for mitochondrial maintenance and longevity. Autophagy 6, doi:11948 [pii] (2010).
- Takahashi, K., Imano, R., Kibe, T., Seimiya, H., Muraatsu, Y., Kawabata, N., Tanaka, G., Matsumoto, Y., Hiromoto, T., Koizumi, Y., Nakazawa, N., Yanagida, M., Yukawa, M., Tsuchiya, E, and Ueno, M. Fission yeast Pot1 and RecQ heloicase are required for efficient chromosome segregation. Mol Cell Biol 31, 495-506 (2011).
- Shiroiwa, Y., Hayashi, T., Fujita, Y., Villar-Briones, A., Ikai, N., Takeda, K., Ebe, M., and Yanagida, M. Mis17 is a Regulatory Module of the Mis6-Mal2-Sim4 Centromere Complex That Is Required for the Recruitment of CenH3/CENP-A in Fission Yeast. PLoS One 6, e17761 (2011).
- Pluskal, T., Hayashi, T., Saitoh, S., Fujisawa, A., and Yanagida, M. Specific biomarkers for stochastic division patterns and starvation-induced quiescence under limited glucose levels in fission yeast. FEBS J, doi: 10.1111/j.1742-4658.2011.08050.x. (2011).
- Pluskal, T., Castillo, S., Villar-Briones, A., and Orešič, M. . MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinformatics 11, doi: 10.1186/1471-2105-11-395 (2010).
- Kiyomitsu, T., Murakami, H., and Yanagida, M. Protein interaction domain mapping of human kinetochore protein Blinkin reveals a consensus motif for bining of spindle assembly checjpoint protins Bub1 and BubR1. Mol Cell Biol 5, 998-1011 (2011).
4.2 Books and other one-time publications
Nothing to be reported4.3 Oral and Poster Presentations
- Yanagida, M. How are non-dividing (quiescent) cells able to keep alive?, Research Workshop FANTASY, Kamakura Prince Hotel, Kamakura, Japan, Feb 5, 2011
- Yanagida, M. Cell Division and Metabolism of Fission Yeast S.pombe, invited seminar at university, Comenius University of Brastilava, Brastilava, Slovakia Jan 21, 2011
- Yanagida, M. ”Metabolic control of chromosome dynamics and cell division" invited seminar at university, Research Institute of Molecular Pathology, Vienna, Austria, Jan 20, 2011
- Yanagida, M. Control of Chromosome Dynamics by Condensin and Metabolism, FEBS Advanced Lecture Course "Trends in Genetics:Genomic Instability and Pathway of Response", Yerevan, Armenia, Feb 20-26, 2011
- Yanagida, M. RNA and one on Development, The EMBO members' workshop, Heidelberg, Germany, Oct 27-29, 2010
- Yanagida, M. Getting Chromosomes Apart, Yeast Genetics & Molecular Biology Meeting, University of British Columbia, Vancouver, Canada, Jul 27-31, 2010
- Yanagida, M. The G-zero project: How fission yeast survive starvation, Cancer Research UK Seminar, Cancer Research UK, London, England, Jun 24, 2010
- Yanagida, M. On clearing mitosis, EMBO Workshop Chromosome Segregation & Aneuploidy, The Surgeons Hall, The Royal College of Surgeons of Edinburgh, Edinburgh, England, Jun 23-24, 2010
- Yanagida, M. Metabolic Control of Quiescence, Aging and Lifespan in Fission Yeast, IGMM Distinguished Seminars, Institute of Genetics and Molecular Medicine, Edinburgh, England, Jun 17, 2010
- Yanagida, M. Condensin and Clearing Mitosis, University seminar, University Heidelberg, Germany, Jun 16, 2010
- Yanagida, M. Mostly DNA, a Bit of Glucose, and the Next 45 Minutes, The Hunt Symposium, London, UK, Dec 9, 2010
- Yanagida, M. Cell Division and Metabolism, Cellular Dynamics and Chemical Biology, Hefei, China, Nov 19-22, 2010
- Yanagida, M. Genetic control of reversible cellular quiescence, inStem Annual Symposium, Bangalore, India, Nov 12-14, 2010
- Tomas Pluskal, T. H., Shigeaki Saitoh, Asuka Fujisawa, and Mitsuhiro Yanagida. Minimal glucose requirements for cellular happiness, OIST Junior Research Retreat, Okinawa, Japan, Nov 2, 2010
- Takeshi Hayashi, T. P., Nobuyasu Ikai, Mitsuko Hatanaka, Mitsuhiro Yanagida. Analysis of Tel2-Tti1 complex regulating TOR pathways in fission yeast, BMB2010, Kobe, Japan, Dec 7-10, 2010
- Takeda, K. The cooperation of proteolytic machineries required for the maintenance of mitochondria and cellular lifspan in fission yeast G0 phase, JST CREST 3rd symposium), University of Tokyo, Japan, Oct 21, 2010
- Takahiro Nakamura, T. P., Yukinobu Nakaseko, and Mitsuhiro Yanagida. CoA biosynthesis pathway is regulated by Sir2 and affects chromosome segregation through protein acetylation, BMB2010, Kobe, Japan, Nov 7-10, 2010
- Pluskal, T., Nakamura, T., Villar-Briones, A., Takeda K., Yanagida, M. Fission yeast metabolites implicated in stress response, nutrient starvation and longevity, Metabolomics 2010, Amsterdam, Jun 27-Jul 1, 2010
- Pluskal, T., Nakamura, T., Villar-Briones, A., Yanagida, M. Fission yeast metabolites implicated in stress response, nutrient starvation and longevity, JST CREST 3rd symposium), University of Tokyo, Tokyo, Japan, Oct 21, 2010
- Nakazawa, N. Condensin phosphorylated by aurora B-like Ark1 is continuously required till telophase in the mode distinct from Top2, The 9th international student seminar, Kyoto University, Kyoto, Japan, Mar 7, 2011
- Nakamura, T. CoA biosynthesis pathway is regulated by Sir2 and affects chromosome segregation through protein acetylation, JST CREST 3rd symposium), University of Tokyo, Tokyo, Japan, Oct 21, 2010
- Kojiro Takeda, T. Y., Sakura KIkuchi, Koji Nagao, Aya KOkubu, Tomas Pluskal, Alejandro Villar-Briones, and Mitsuhiro Yanagida. Cooperative roles of the proteasome and autophagy for mitochondrial maintenance and lifespan in the quiescent phase in fission yeast, BMB2010, Kobe, Japan, Dec 7-10, 2010
- Kenichi Sajiki, Y. T., and Mitsuhiro Yanagida. Screening of essential genes for quiescence using S.pombe deletion mutant library sets, BMB2010, Kobe, Japan, Dec 7-10, 2010
- Kenichi Sajiki, Y. T., and Mitsuhiro Yanagida. Screening of essential genes for quiescence using S.pombe deletion mutant library sets, BMB2010, Kobe, Japan, Dec 7-10, 2010
- Ikai, N. In Vitoro Kinase Assay of Two Tor Complexes in S.Pombe and Construction of a Temperature-Sensitive Tor1 Mutant, JST CREST 3rd symposium, University of Tokyo, Tokyo, Japan, Oct 21, 2010
- Hayashi, T., Hatanaka, M., Kanoh, J., Ikai, N., and Yanagida, M. Tel2-Tti1 complex regulates TOR pathways in fission yeast, JST CREST 3rd symposium, University of Tokyo, Tokyo, Japan, Oct 21, 2010
5. Intellectual Property Rights and Other Specific Achievements
Nothing to be reported6. Meetings and Events
Nothing to be reported