



Developmental Signalling Unit
Principal Investigator: Mary Ann Price
Research Theme: Genetic, biochemical, and cell biological studies on the mechanism of Hedgehog signalling

Abstract
Hedgehog (Hh) signalling plays many roles in cell and tissue specification, patterning, and stem cell regulation during the development of many multi-cellular animals. In Drosophila, where it was first discovered, Hh is involved in such diverse roles as embryonic segmentation, appendage development, and the maintenance of stem cell like compartments in the ovary and lymph gland. In vertebrates, Hh homologues have been shown to have important roles in specification of neurons in the developing spinal cord and brain, the anterior-posterior patterning of the limbs, and in early thymocyte (T lymphocyte precursor) development in the thymus.
The Hh pathway, named for the ligand Hh, operates primarily by causing changes in target gene expression in the cells that bind to Hh. In Drosophila, the effects of Hh on transcription are mediated by the Gli family transcription factor Cubitus interruptus (Ci). In the absence of Hh signalling, full-length Ci (Ci-155) is processed to a smaller repressor form (Ci-75). Hh signalling inhibits this processing and converts Ci-155 to a more potent transcriptional activator. Thus, this Hh-regulated switch in Ci activity is important to determine the gene expression in cells and ultimately their identity.
In the wing imaginal disc, the precursor to the adult wing, Hh is expressed in posterior (P) compartment cells and signals to anterior (A) compartment cells, which express Ci and Ptc, the Hh receptor. Hh secreted by P cells binds to A cells near the A/P border, leading to an increase in full-length Ci levels and in target gene expression in these A cells. One such target is decapentaplegic (dpp), a TGFb family member involved in the growth and patterning of the wing disc (Figure 1).
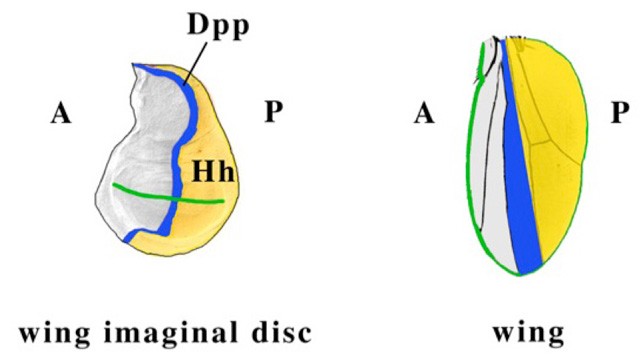
Figure 1. A schematic of the adult wing (right) and wing imaginal disc (left), the larval structure that becomes the adult wing. Hh is expressed in the posterior (P) compartment cells (shown in yellow); Ci and Ptc are expressed in the anterior (A) compartment cells. Hh diffuses to A cells near the A/P border, binds to Ptc, leading to a block in Ci processing and thus high levels of Ci-155 at the border, which in turn leads to upregulation of target genes, such as dpp (blue). The green line marks the margin of the wing and the presumptive wing margin in the wing imaginal disc.
In the larval lymph gland, Hh is expressed in the PSC (posterior signalling center) and is involved in maintaining a stem-cell like compartment (termed the medullary zone), preventing these cells from prematurely differentiating into hemocytes (insect blood cells). As in the wing disc, full-length Ci levels are highest near the source of Hh (Figure 2).
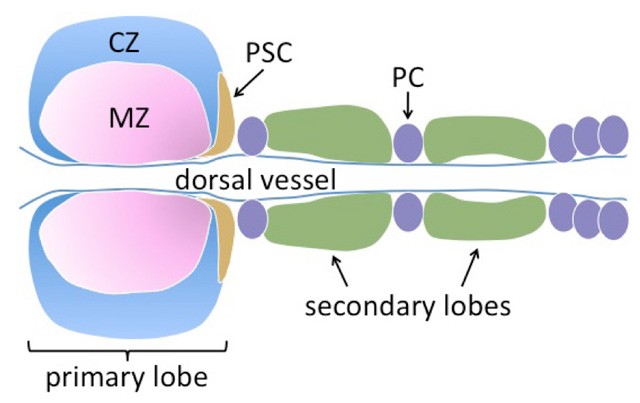
Figure 2. A schematic of the larval lymph gland. This tissue is derived from embryonic mesodermal cells and flanks both sides of the dorsal vessel (the insect heart). A specialized group of cells termed the posterior signalling center (PSC) expresses signals that maintain cells in the medullary zone (MZ) in a stem-cell like, undifferentiated state. These cells give rise to differentiated hemocytes in the cortical zone (CZ). Hh is expressed in the PSC. Ci (pink) and Ptc are expressed in the MZ. Ci-155 levels are highest near the source of Hh in the PSC. Pericardial cells are shown as purple ovals.
The current model for Ci processing is as follows. In the absence of Hh signalling, Cos2, a kinesin-like molecule, binds Ci and three kinases (PKA, CK1, and GSK3). Ci is phosphorylated by the three kinases, followed by binding of Slimb, part of an E3 ubiquitin ligase complex, to phosphorylated Ci. Ubiquitinated Ci then becomes the target of partial degradation by the proteasome. In the presence of Hh, Ci and the kinases dissociate from Cos2, hypo-phosphorylated Ci-155 accumulates and is not a target of proteasomal processing. While the basics of Ci processing are understood, there are still many outstanding questions, such as why is Ci only partially degraded by the proteasome, which normally completely degrades its targets, and does Hh do more than cause dissociation of the Cos2 complex in order to inhibit Ci processing.
My lab is primarily interested in how Hh signalling regulates Ci activity and processing. We are also interested in the role of Hh signalling in T-cell differentiation and proliferation. To this end, we have undertaken the following projects:
- identification of novel genes involved in Ci processing using a genome wide double-stranded RNA interference (dsRNAi) screen, and characterization of these genes in the wing and lymph gland
- characterization of the subcellular localization of Ci and other Hh signalling components and the regulation of this localization by Hh
- production and purification of Hh signalling components for biochemical and structural assays
- elucidation of the potential role of Hh signalling in T-cell proliferation and differentiation
1. Staff
- Ms. Chika Azama, Research Administrator/Secretary
- Dr. Michiko Arai, Technician
- Dr. Satoshi Hasegawa, Technician/Post-doc
- Dr. Spencer Spratt, Post-doc
- Dr. Olga Elisseeva, Post-doc
- Dr. Umesh Gangishetti, Post-doc (started Nov. 1, 2010)
- Dr. Yasuji Kido, Post-doc (shared with Samatey unit, started Nov. 1, 2010)
- Dr. Marco Tsui, Post-doc (started Dec. 15, 2010)
- Dr. Ingo Lehmann, Post-doc (left July 31, 2010)
- Dr. Anri Itoh, Researcher (July 14, 2010 – Jan. 28, 2011)
2. Collaborations
Nothing to report.3. Activities and Findings
3.1 RNAi Screen
In order to identify new genes required for Ci processing in the absence of Hh, we previously conducted a genome-wide RNAi screen in Kc cells, a Drosophila embryonic hemocyte line. Briefly, Ci is tagged at the N-terminal end with firefly luciferase and at the C-terminal end by Renilla luciferase and expressed in a stable cell line. The activity of both these enzymes can be measured in a single sample. Because the Renilla luciferase is degraded along with the C-terminal end of Ci during Ci-75 formation, the ratio of Renilla luciferase to firefly luciferase measures the amount of Ci-75 formation. Our screen identified ~250 dsRNAs that either enhanced or blocked Ci processing. We are continuing to follow-up on these candidate genes. This first involved retesting all “hits” in the screen assay; about half of our candidates proved to be false positives. All remaining hits are being tested in the following ways: (1) using a more direct in vitro assay for Ci-75 formation, an IP-Western blot, in which both Ci-155 and Ci-75 are visualized on the same gel, (2) using the initial Renilla/firefly ratio assay with a validation amplicon, a dsRNA targeted to different region of the gene from the original screen amplicon, and (3) testing for an effect in Drosophila in the wing imaginal disc and in the lymph gland. Because some of our strongest hits in the Kc cell assays did not show an effect or showed only a weak effect in wing discs, we have begun testing these candidates in the larval lymph gland, with initial results looking promising (Figure 3).
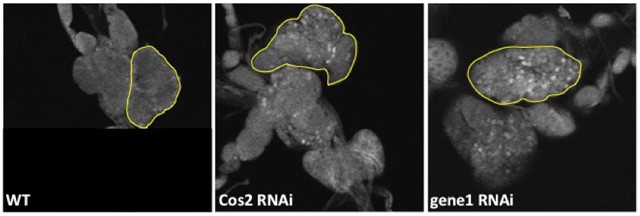
Figure 3. Lymph glands stained with an antibody for full-length Ci. Lymph glands are wild-type (left panel), expressing a Cos2 RNAi (center panel) or expressing an RNAi to a novel gene found in our screen (right panel). The lymph glands expressing Cos2 RNAi and RNAi for the novel gene show more cells with a high level of full-length Ci relative to the wild-type lymph glands. The yellow line outlines one primary lobe of the lymph gland. RNAi expression is driven by domeless-GAL4, which is expressed in the medullary zone.
We have recently received external funding to conduct a second RNAi screen designed to identify genes required for the Hh pathway to block Ci processing.
3.2 Subcellular localization of Hh signalling components
Since Ci has been suggested to partially localize to intracellular vesicles and our previous results confirmed a punctate pattern for cytoplamsic Ci in wing imaginal disc cells (Figure 4), we are attempting to identify which type of vesicles Ci localizes to. We are also trying to determine whether this localization is important for Ci-75 formation and/or Ci regulation by Hh signalling.
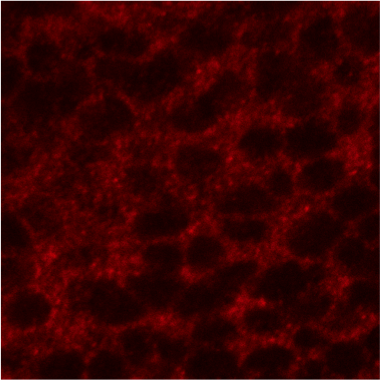
Figure 4. Anterior compartment of a wing imaginal disc stained with 2A1 antibody (red), which recognizes only Ci-155. Ci-155 shows a punctate pattern of staining on top of a lighter diffuse signal in the cytoplasm. These puncta may represent intracellular vesicles, such as endosomes or lysosomes. The large circular black regions are nuclei. (Ci-155 only shows nuclear staining when nuclear export is blocked.)
3.3 Expression and purification of Hh signalling components for biochemical and structural studies
In collaboration with Fadel Samatey’s unit, we are utilizing a number of protein expression systems for making Hh signalling components, which we will purify and use in biochemical and structural assays.
3.4 Hh signalling in lymphocytes
We are examining whether Hh signalling has a role in modulating lymphocyte differentiation by studying both adult peripheral blood lymphocytes, which can differentiate into memory cells and effector cells, and lymphocyte precursors in umbilical cord blood. In both cases, we find the components necessary for Hh signalling present in the cells (in particularly high levels in precursor cells) and in the case of adult lymphocytes, the cells express Hh target genes in response to activation of the pathway.
4. Publications
4.1 Journals
Nothing to report.4.2 Books and other one-time publications
Nothing to report.4.3 Oral and Poster Presentations
Price MA. Mechanism of the Partial Proteolysis of Cubitus interruptus, HHMI summer undergraduate research program University of Alabama, Tuscaloosa, AL, USA, Jun 02, 2010
Price MA. Invertebrate Embryology and Patterning, Developmental Neuroscience Course (DNC), OIST, Onna-son, Okinawa, Jul 12 - 22, 2010
Price MA, W. Y., Arai M, Hasegawa S, Spratt SJ. The mechanism of Ci repressor formation, EMBO Workshop Hedgehog Signaling: From developmental biology to anti-cancer drugs, St. Jean, France, Mar 27-31, 2010
Spencer, S. A genome-wide search for novel Hedgehog signalling regulators involved in Cubitus interruptus (Ci) processing, OIST Junior Researcher Retreat, Nov 2, 20105. Intellectual Property Rights and Other Specific Achievements
Nothing to report.6. Meetings and Events
6.1 Workshop
- Title: OIST Developmental Neurobiology Course
- Dates: July 12-22, 2010
- Co-organizers:
- Mary-Ann Price (OIST)
- Davie Van Vactor (Harvard/OIST)
- Akinao Nose (University of Tokyo)
- Ichiro Masai (OIST)
- Robert Baughman (OIST)
- Speakers:
- Yoshiki Sasai (Riken CDB)
- Li-Huei Tsai (MIT)
- Masayuki Miura (University of Tokyo)
- Frank Gertler (MIT)
- Elke Stein (Yale)
- Tadashi Uemura (Kyoto University)
- Kang Shen (Stanford)
- Morgan Sheng (Genentech)
- Hitoshi Okamoto (Riken BSI)
- John Flanagan (Harvard)
- Nirao Shah (UCSF)
- Hideyuki Okano (Keio University)
- Alex Whitworth (University of Sheffield)
- Thomas Walz (Harvard Medical School)
7. Others
OIST Open Campus (co-organizer, 2 exhibits), Nov. 2010, “DNA: the Blueprint of Life (banana DNA extraction)” and “Colors and Science (red cabbage litmus and candy chromatography)”
Onna/OIST Children’s School of Science (course organizer for 3rd and 4th grade course “Oishii Kagaku/Delicious Science”)