



Computational Neuroscience Unit
Principal Investigator: Erik De Schutter
Research Theme: Modeling Cellular and Molecular Mechanisms of Neural Information Processing

Abstract
We use computational, data-driven methods to study how neurons and microcircuits in the brain operate. We are interested in the interaction between fundamental properties like morphology or excitability and neural functions like information processing or learning. Most of our models concern the cerebellum as this brain structure has a relatively simple anatomy and the physiology of its main neurons has been studied extensively, allowing for detailed modeling at many different levels of complexity.
In FY2010 significant progress has been made on several research projects. Highlights include the layered approach used in NineML, based on theoretical work in the unit. Further development of STEPS includes the option to import SBML models. We performed a deep analysis of how noise affects the signaling pathways involved in induction of cerebellar LTD. We developed efficient methods to simulate buffered calcium diffusion in large neuron models and started modeling the chloride metabolism of Purkinje cells. We are analyzing the relationship between neighboring trees in forests of Purkinje cell dendritic trees and developed a simple model for the growth of dendritic bifurcations. We showed that gap junctions between Golgi cells are less important in generating cerebellar oscillations than previously reported. We have started analyzing complex spike data from monkey cerebellar recordings.
1. Staff
General services and neuroinformatics
Technical Staff: Ivan Raikov
Research Administrator / Secretary: Tsuyuki Nakabayashi
Molecular modelling
Researchers: Gabriela Antunes, Weiliang Chen
Technical Staff: Iain Hepburn
Cellular modelling
Researchers: Sungho Hong, Shiwei Huang, Yihwa Kim
Techmical Staffs: Haroon Anwar, Werner Van Geit (till November 2010)
Research Assistant / Graduate Student: Hermina Nedelescu, Pascal Warnaar
Network modeling
Researchers: Fabio M.S. de Souza, Mario Negrello, Rodrigo Publio (till October 2010)
Research Assistant / Graduate Student: Shyam K. Sudhakar
2. Collaborations
- Type of collaboration: Scientific collaboration and graduate program
- Researchers:
- Professor M. Giugliano, University of Antwerp, Belgium
- Professor D. Snyders, University of Antwerp, Belgium
- Dr. Q. Robberecht, University of Antwerp, Belgium
- Dr. K. Tahon, University of Antwerp, Belgium
- Dr. K. Veys, University of Antwerp, Belgium
- Type of collaboration: Scientific collaboration
- Researchers:
- Professor M.L. Linne, Tampere University of Technology, Finland
- Dr. K. Hituri, Tampere University of Technology, Finland
- Type of collaboration: Scientific collaboration
- Researchers:
- Professor H.P. Thier, University of Tübingen, Germany
- Dr. A. Ignashchenkova, University of Tübingen, Germany
- Dr. M. Junker, University of Tübingen, Germany
- Dr. A. Schmigdlin, University of Tübingen, Germany
- Type of collaboration: Scientific collaboration
- Researchers:
- Professor G.J. Augustine, Korea Institute for Science and Technology (KIST), Korea
- Professor K. Tanaka, Korea Institute for Science and Technology (KIST), Korea
- Dr. J. Kim, Korea Institute for Science and Technology (KIST), Korea
- Type of collaboration: Scientific collaboration
- Researchers:
- Professor F. Santamaria, University Texas, San Antonio, United States of America
- Type of collaboration: Scientific collaboration
- Researchers:
- Professor M. Ellisman, University California, San Diego, United States of America
- Professor M. Martone, University California, San Diego, United States of America
- Dr. L. Fong, University California, San Diego, United States of America
- Type of collaboration: Scientific collaboration
- Researchers:
- Professor C.F. Valenzuela, University of New Mexico, United States of America
- Type of collaboration: Scientific collaboration
- Researchers:
- Professor S.A. Prescott, University of Pittsburgh, United States of America
- Dr. S. Ratté, University of Pittsburgh, United States of America
- Type of collaboration: Scientific collaboration
- Researchers:
- Professor M. Häusser, University College London, United Kingdom
- Dr. H. Cuntz, University College London, United Kingdom
- Dr. A. Roth, University College London, United Kingdom
- Dr. A. Watt, University College London, United Kingdom
- Type of collaboration: Scientific collaboration
- Researchers:
- Professor M. Jusoola, University of Sheffield, United Kingdom
- Dr. Z. Song, University of Sheffield, United Kingdom
3. Activities and Findings
3.1 Neuroinformatics standards
3.1.1 A standard language for multiscale modeling in neuroscience
The International Neuroinformatics Coordinating Facility (INCF) has assembled a task force of simulator developers to propose a declarative computer language for descriptions of large-scale neuronal networks. The name of the proposed language is "Network Interchange for Neuroscience Modeling Language" (NineML). Its initial focus is restricted to point neuron models and uses a layered approach as advocated by our previous work (Raikov and De Schutter, 2011).
As part of the ongoing development effort on NineML, we have implemented code generation software for the diagram-based notation used by NineML for describing hybrid dynamics, which is capable of generating code for several interpreted and compiled programming languages. NineML uses set operations to construct composite neuronal populations and connectivity patterns. In order to describe populations, we have implemented a software library for compact representation of very large sets of neuronal populations. This library is complemented by a library of graph functions that can express connectivity between large populations compactly.
In addition, NineML has as design aims to provide tool support for explicit declarative mathematical definitions of spiking neuronal network models in a simulator independent manner, and to allow addition of new model descriptions without modification of the previous structure and organization of the language. To meet these goals, we have developed an initial prototype of a NineML model description interpreter, which integrates the different aspects of NineML in order to allow the construction of complete models, from neuronal and synaptic dynamics to complete networks. The interpreter is capable of reading specialized syntax for NineML models, as well as a preliminary XML syntax, and can invoke the code generation functions for the currently supported programming languages and neuronal simulators. The interpreter provides a consistent and unified user interface to the different model description functionality of NineML.
3.2 Molecular mechanisms of synaptic plasticity
3.2.1 STEPS software development
STEPS (Wils and De Schutter, 2009) is a software system for simulation of biological reaction-diffusion systems in well-mixed systems or complex 3D morphologies, such as the signaling pathways involved in synaptic plasticity.
Computational efficiency has always been one of the major focuses in the development of STEPS, since SSA simulations are commonly time consuming. Several update versions of STEPS were released after extensive profiling and reimplementation, reducing simulation time significantly. We implemented a recent SSA solution with lower computational complexity (Slepoy et al., 2008).
Improvement was made in the mesh preparation for STEPS simulations. Traditionally tetrahedral meshes were constructed by combinations of predefined geometry primitives, such as cylinders and spheres (Santamaria et al., 2006). Although this method provides fast solution for mesh generation, the lack of biological realism of generated meshes limits their application. We have added the option to generate 3D meshes from the skeleton of more accurate, surface meshes of dendritic sections, which are reconstructed from electron microscopy image series.
3.2.2 Support for SBML in STEPS
The Systems Biology Markup Language (SBML; Hucka et al. 2003) has become a de facto standard for representing computational models of networks of biochemical interactions. The neuronal signaling pathways that STEPS is designed to simulate in a well-mixed context fall into this category. Therefore, support for SBML is an important aspect of STEPS development.
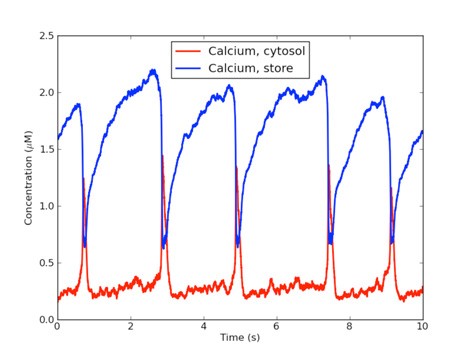
Figure 1: SBML model of calcium oscillations simulated in STEPS. The Goldbeter et al. (1990) model is solved with stochastic methods in femtolitre compartments.
SBML has evolved from the assumption that the simulator will solve differential rate equations, which is somewhat different from the stochastic approach where the underlying maths is based on one fundamental, unchanging stochastic reaction constant. This presented several challenges when developing SBML support, particularly the need to read, store and solve mathematical expressions provided in MathML format. STEPS now provides structures to represent the mathematical expressions in nested Python sequences, along with functions for solving them. Such expressions may then be saved and updated during simulation, which has allowed support for many important features of SBML such as Events and Rules. Also, by providing methods to compare the mathematics specifically for a reaction 'speed' to the form of a fundamental reaction, we can separate reactions into those that can be represented as an ordinary reaction in STEPS and those of which the form differs, and so are solved by an approximate method. This MathML support means that STEPS represents all reactions correctly. Any stochastic simulator that ignored such mathematical expressions would represent the majority of SBML models incorrectly.
SBML support in STEPS has been tested against the extensive SBML 'Test-Suite' and against many models from the Biomodels database (Li et al. 2010). 71% of the first 900 Test-Suite models are supported, and the majority of the failed imports are unphysical models of 0-dimensional or 1-dimensional compartments or incomplete models with no chemical species present. Figure 1 shows an example stochastic simulation of an SBML model of signal-induced calcium oscillations (Goldbeter et al. ,1990) in STEPS.
3.2.3 Stochastic modeling of induction of cerebellar long-term depression
Many cellular processes involve small number of molecules and are susceptible to undergo stochastic fluctuations in their level of activity. Among these processes is the induction of cerebellar long-term depression (LTD), a long-lasting decrease in the efficacy of the excitatory synaptic transmission between granule cells and Purkinje cells, expressed as a reduction in the number of synaptic AMPA receptors (AMPARs). Cerebellar LTD can be induced through elevations in the postsynaptic intracellular calcium ions concentration ([Ca2+]i), which leads to the activation of a positive feedback loop fundamental for LTD occurrence that includes the Ca2+-dependent protein kinase Cα (PKC) and the cytosolic phospholipase A2 (cPLA2), both activated directly by [Ca2+]i, and the mitogen-activated protein kinase (MAPK) pathway, that is activated by PKC. The persistent activation of PKC regulates the trafficking of AMPARs, leading to the synaptic depression. We developed a stochastic model of the signaling network and mechanisms of AMPAR trafficking involved in LTD, and showed that the network activity switches between two discrete stable states. Fluctuations in the signaling network activity lead to the probabilistic induction of LTD, which gives rise to the macroscopic sigmoidal relationship between LTD and the concentration of the triggering signal observed experimentally. Increasing the biochemical population size modifies quantitatively and qualitatively its macroscopic behavior (Figure 2), which is caused by the reduction of the stochastic fluctuations of the signaling network.
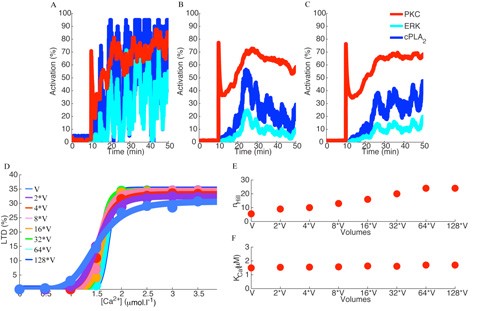
Figure 2: Fluctuations in the activity of the signaling network are controlled by the biochemical population size. Increasing the biochemical population size, which was done by increasing the volume of the biochemical compartment where the reactions take place, reduces the stochastic fluctuations of the model (A – original population, B – population 32 times larger, C – population 64 times larger). D: Alterations in the population size of the model changes the sigmoidal relationship of LTD as a function of the input signal, potentiating the bistability observed during LTD induction. E: The alterations in the macroscopic bistability of LTD induction can be measured by the hill coefficient (nHill) of the sigmoid curves, which increase as a function of the population size represented here by the volume of the biochemical compartment. F: the [Ca2+]i requirement for LTD induction (KCa) is independent of the population size.
3.3 Cellular mechanisms regulating firing and synaptic properties of neurons
3.3.1 Improvements of a Purkinje neuron model
We have recently obtained voltage clamp data of the P-type calcium current recorded in Dr. Indira Raman's lab (Northwestern University). The P-type calcium channel is present through all parts of the Purkinje neuron including the axon, and plays an important role in excitability not only by itself but also by contributing to activation of the calcium-activated potassium channels. One of the main issues of computational modeling this channel is that it both expresses calcium-induced facilitation (CIF) and inactivation (CII), but full under-standing of the biophysical mechanisms causing them has yet not been established (Dunlap, 2007). CIF usually expresses significantly when the channel is only partially activated. Since the baseline membrane potential of the dendrites is around this partial activation range, it is important to have a model with CIF to test its influence on the entire Purkinje neuron model.
We adopted the suggested mechanism by Chaudhuri et al. (2007) and implemented it in our P-type channel model, and it successfully reproduced the CIF in the experimental data. We are currently investigating the effects of CIF in the full Purkinje neuron model.
3.3.2 Modeling detailed calcium dynamics in the Purkinje cell dendrite
Intracellular Ca2+ concentration plays a crucial role in the physiological interaction between Ca2+ channels and Ca2+-activated K+ channels. We have investigated the role of different Ca2+ buffering mechanisms in simulating this physiological interplay (Anwar et al., 2011). The exponential decaying pool used in most multi-compartmental neuron models could not simulate characteristic Purkinje cell Ca2+ spikes. The reason is its short relaxation time which can approximate fast transients and therefore activate a BK-type channel, but fails for the small conductance (SK) channel that activates slowly and requires different Ca2+ dynamics. A simple and effective solution may be to use two Ca2+ pools respectively for the fast and slow transient. Alternatively one could use a detailed model of Ca2+ dynamics (Schmidt et al., 2003) including Ca2+ diffusion, diffusible parvalbumin and calbindin buffers and pumps. Unfortunately simulating diffusion is very computation intensive and would make simulations of a multi-compartmental model prohibitively slow. Therefore we developed a Diffusion Compensation Method (DCM) that replaces diffusion of Ca2+ and buffers with a DCM buffer (Anwar et al., 2011).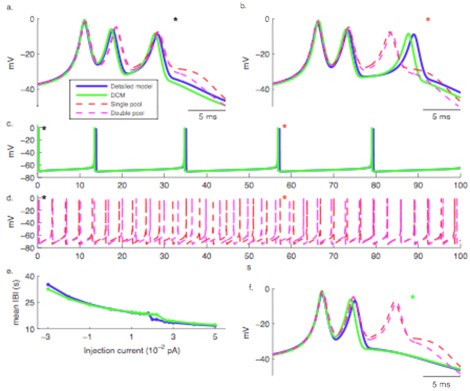
Figure 3: Dendritic Ca2+ spikes generated using different Ca2+ buffering models. Spikes are aligned at the peak of the first spikelets in a, b and f. a. First burst of Ca2+ spikes, b. Burst of Ca2+ spikes around 57 s, c and d, Spontaneous Ca2+ spike bursting over 100 s, e. Inter burst interval (IBI) as a function of current injection, f. Burst of Ca2+ spikes around 57 s with injection of 0.004 pA current. (Anwar et al., 2011).
When dendritic Ca2+ spikes with multiple spikelets comparable to those observed in vitro were simulated using different Ca2+ buffering models (Figure 3a-b), the spikes generated using pool based models were different from the spikes generated using the other buffering models. Differences existed in spike width, spike height and afterhyperpolarization and lead to completely different behavior of the models over longer time scales (Figure 3c-e). On the other hand, the Ca2+ spikes generated using DCM were similar to those generated using detailed model.
While using detailed Ca2+ dynamics models with diffusion in a detailed morphology of a Purkinje cell, we discovered large gradients of Ca2+ levels in neighboring segments with different diameters (Figure 4). The peak Ca2+ concentration showed a close to linearly inverse relationship to the diameter of the compartment. This effect was present also when 3D diffusion was simulated using STEPS. We deem such pronounced gradients of Ca2+ within the dendrite as unphysiological, leading to the paradoxical effect that more realistic models (buffered Ca2+ diffusion) produce unrealistic gradients compared to the simpler pool models which are relatively insensitive to diameter. We are exploring possible regulatory mechanisms to compensate for this effect in neurons.
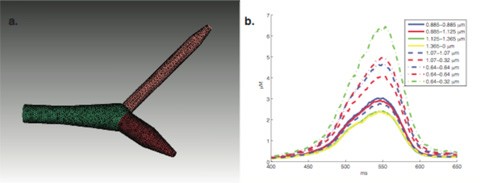
Figure 4: Detailed Ca2+ buffering models cause a strong dependence on dendritic diameter. Ca2+ transients shown in panel b are generated using uniform channel densities in a piece of dendrite with varying diameter as shown in panel a (STEPS simulation).
3.3.3 Modeling chloride dynamics in Purkinje cells
Regulation of intracellular Cl- concentration in neurons is important for synaptic inhibition. The molecular mechanisms underlying this regulation are not well understood. Biophysical properties of many Cl- channels and transporters are examined in heterogeneous expression systems and their physiological role determined from gene expression studies – both of which do not provide information on the dynamics of Cl- regulation. However, these studies can be used to construct a dynamical model of Cl- regulation.
Cl- concentration in Purkinje cells may be regulated by two classes of transporters – the Potassium Chloride Co-transporters (KCCs) and the Excitatory Amino Acid Transporters (EAATs). KCCs utilize the ion gradient of K+ to remove intracellular Cl- (Blaesse et al., 2009) and is co-localized with GABAaRs. EAATs utilize the ion gradients of Na+, H+ and K+ to exclude glutamate from the synapse (Zerangue and Kavanaugh, 1996). In addition to their transport function, the binding of Na+ and glutamate to the transporter activate an uncoupled channel within the transporter, permeant to monovalent anions (Fairman et al., 1995). Purkinje cell EAAT current is predominantly an anion current as replacing Cl- with a more permeant anion greatly enhances the current amplitude (Otis et al., 1997). This current may contribute to Cl- regulation, particularly in low intracellular volume compartments (e.g. spines) where the Cl- gradient can fluctuate rapidly.
Detailed models of KCC2 and Purkinje cell EAATs were constructed and incorporated into a biophysically and morphologically realistic model. The model will be used to explore conditions at which KCC2 and Pn EAATs can alter Cl- reversal potential and IPSC amplitudes.
3.3.4. Efficient estimation of the phase response curve via compressive sensing
The phase response curve (PRC) is an important characterization of biological oscillators such as neurons. However, estimation of the PRC from the experimental data has posed a problem due to a large mismatch between the dynamical time scales of the stimuli injected during the experiments and neuronal intrinsic mechanisms (Torben-Nielsen et al, 2010). Recently, we developed a new method to estimate the PRC based on a recent breakthrough in information theory, called compressive sensing (Baraniuk, 2007). The method shows good results with surrogate data. It will be applied to PRCs from cerebellar Golgi cells, recorded at the University of Antwerp.
3.3.5 Modeling dendritic growth with simple models
How neurons attain their specific shapes has attracted attention over centuries. The dendritic shape determines the type of input a cell receives. Cell-internal as well as external mechanisms are known to contribute in determining the characteristic morphology of each cell type. We try to identify the contributions of such internal and external mechanisms to growth using modeling approaches.
In order to assess the morphological property of each neuron type, we have used dendritic bifurcations as the unit of study due to their abundance in number and the fact that only three measures are sufficient to completely account for their geometrical properties. Since within each neuron type variability exists in the shape, single cell reconstruction data was pooled from the Neuromorpho.org database (Ascoli et al. 2007) and statistical properties of the bifurcation shapes of neurons were studied.
A growth model that was based only on internal mechanisms guiding development was tested and found to match the characteristic bifurcation shapes of most neurons.
3.4 Information processing in the olivocerebellar system
3.4.1 Morphological variability in populations of neighboring Purkinje cells
For decades anatomists have primarily relied on reconstructing dendritic arbors at the rate of only a single or a few sparsely located cells at a time. Such studies have generated only a partial understanding of a network's cytoarchitecture and underlying neuronal plasticity throughout life.
To gain a more comprehensive insight into the architecture of anatomical networks, we have commenced to explore the nature of structural changes occurring among neighboring Purkinje cells as a function of neuronal growth. In collaboration with Alanna Watt and Hermann Cuntz of the University College London, we study large forest populations of Purkinje cells. We have been able to investigate the morphological changes at different neuronal growth stages of hundreds of neurons by combining transgenic mouse technology and microscopy with computational anatomy. Figure 5 demonstrates a small sample of a reconstructed Purkinje cell forest from the cerebellum of a Green Fluorescent Protein (GFP) expressing transgenic mouse in which specifically Purkinje cells are labeled by GFP. We have made further progress by employing Brainbow technology as a tool in our research to facilitate differentiation of neighboring Purkinje cells. Thus far, our results suggest that there is considerably more organizational and morphological variability among neighboring Purkinje cells in the network then previously thought.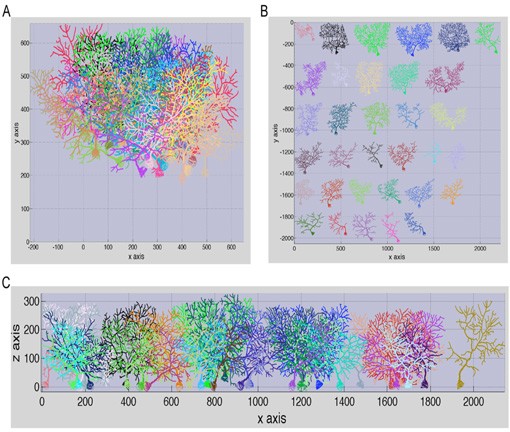
Figure 5: Sample of anatomical reconstructions from the cerebellum of a young GFP transgenic mouse. A: Intact forest spanning ~211 X 211 X 134 μm and containing 33 Purkinje cells. False colors are arbitrary to facilitate visualization of neighboring arbors. B: Spread out visualization of individual neurons composing the dense forest in (A). Note the immense diversity among neighboring neurons and their subtrees. C: Illustration of the forest, spread out, in the x-z view. All images were plotted with the TREES toolbox.
3.4.2 Predicted effect of ethanol on granular layer oscillations
A network model of cerebellar cortex has been developed to simulate the actions of ethanol on cerebellar function. This is based on a previously constructed neural network model of the cerebellar granular cell layer (Simões de Souza and De Schutter, 2011), and work that studies the excitatory action of ethanol on single cerebellar Golgi Cells by inhibiting the Na+/K+ ATPase (Botta et al., 2010).
We first used the neural network model to explore the unclear role of gap junctions between Golgi cells on the generation of cerebellar electrical oscillations. We discovered that gap junctions only increase the robustness of oscillations that are primarily driven by a synaptic feedback loop between Golgi and granule cells (Simões de Souza and De Schutter, 2011; Figure 6). We are extending these studies to investigate the effects of ethanol on the granular layer oscillations.
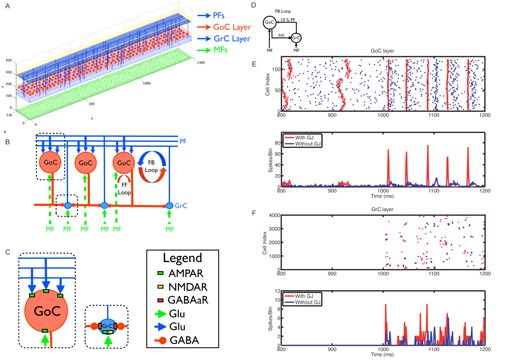
Figure 6: Neural network topology and effect of gap junctions on network in the oscillations. A The spatial location of neurons in their two-dimensional layers is shown. Only 1% of the granule cells and parallel fibers are displayed for better visualization. B Schematic diagram illustrating the connectivity between the layers. C Inset showing the neurotransmitters and synaptic receptors used by each modeled synaptic connection. D The influence of gap junctions between GoCs on the responses of the network in the complete configuration with weak parallel fiber inputs. E Raster plot (top panel) and PSTH (bottom panel) of the Golgi cells with (red) and without (blue) gap junctions. Each dot in the raster plot is a spike. Mossy fiber input with mean firing rate at 5 Hz was turned on at 1000 ms. F Same as (E) but for the granule cells.
3.4.3 Complex effects of the complex spike
Along with other unique architectonic and dynamic particularities, the cerebellum receives one of the most peculiar inputs in the whole brain, the climbing fiber. A single climbing fiber axon lays over the exuberant arbor of the Purkinje neuron as ivy, with hundreds of synaptic contacts producing one of the strongest cell to cell contacts in the whole brain. The climbing fiber action potentials are relatively rare — approximately one per second — and lead the Purkinje neuron to the production of a 'complex spike'. This is a momentous event, which is bound to have significant functional implications for higher functions such as movement coordination and learning.
In collaboration with the Primate Lab of the University of Tübingen, we study the influence of the complex spike measured from the primate cerebellum in both spontaneous and guided oculo-motor movements. We analyze the effects of the complex spike on (1) the local field potentials (Figure 7), (2) simple spike trains and pauses, and (3) on oculo-motor movements. Our results seem to imply an interpretation that is at odds with functions commonly associated with the complex spike, and indicate a more direct impact on behavior than previously believed.
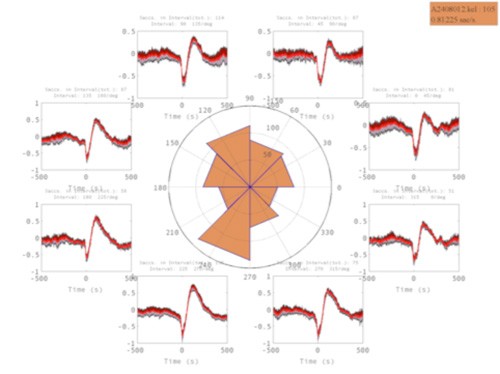
Figure 7: Impact of a saccade to 8 directions on the local field potential in cerebellum.
No thorough attempt has ever been made to describe the variety of complex spikes seen in the cerebellum in vivo. Using the same data sets and employing spectral analysis to subdivide complex spikes we have found multiple forms for different recordings, but also in single recordings a clear variety can be seen. The differences among the complex spikes consist of spikelet number differences and frequency of spikelets differences (Figure 8). Further investigation will focus on functional relevance of the different complex spike waveforms seen.
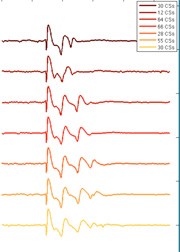
Figure 8: Variability in complex spike shapes in vivo. Data obtained from a single Purkinje cell in monkey cerebellum. Inset shows number of occurrences of each shape in one recording.
3.4.4 Intrinsic heterogeneity of the superchiasmatic nucleus
The superchiasmatic nucleus (SCN) is a pacemaker of the circadian rhythm in mammalian hypothalamus. SCN is often referred as an example of dynamic synchronization because the oscillating neurons, despite manifest heterogeneity in isolation, can generate a coherent rhythm due to intrinsic nonlinearities and mutual coupling. However, the imaging experimental data obtained by our collaborators Jihwan Myung (Kyoto University) and Tohru Takumi (Hiroshima University) show evidence that gene expression in SCN neurons still has some heterogeneity even when the whole slice generates a coherent rhythm. A problem in analyzing this data was that the mean oscillation is so dominating that the typical pairwise correlation is close to one, making it difficult to obtain topographically stable patterns from the observed heterogeneity. We developed a method, based on partial correlation and multi-scale spectral clustering algorithm (Azran and Ghahramani, 2006), to classify the neurons in the imaging data into clusters according to similarities in their oscillating patterns.3.5 References cited
Anwar H, Hong S, De Schutter E (2011) Controlling Ca2+-activated K+ channels with models of Ca2+ buffering in Purkinje cells. Cerebellum: in press. doi: 10.1007/s12311-010-0224-3.
Azran A, Ghahramani Z (2006) Spectral methods for automatic multiscale data clustering. IEEE Computer Society Conference on In Computer Vision and Pattern Recognition. vol 1.Ascoli GA, Donohue DE, Halavi M (2007) Neuromorpho.org: A central resource for neuronal morphologies. J Neurosci 27(35):9247-9251.
Baraniuk RG (2007) Compressive sensing. IEEE Sig Proc Mag 24: 118.
Blaesse, P., Airaksinen, M. S., Rivera, C., and Kaila, K. (2009). Cation-chloride cotransporters and neuronal function. Neuron 61, 820-838.
Botta P., Simoes de Souza F. M., Sangrey T., De Schutter E.D., Valenzuela F. (2010) Alcohol excites cerebellar Golgi cells by inhibiting the Na+/K+ ATPase. Neuropsychopharmacology 35: 1984-1996.
Dunlap K (2007) Calcium channels are models of self-control. J Gen Physiol 129: 379-383.
Chaudhuri D, Issa JB, Yue DT (2007) Elementary mechanisms producing facilitation of Cav2.1 (P/Q-type) channels. J Gen Physiol 129: 385-401.
Goldbeter A, Dupont G, Berridge MJ (1990) Minimal model for signal-induced Ca2+ oscillations and for their frequency encoding through protein phosphorylation. Proc Natl Acad Sci USA 87:1461-1465.
Fairman, W. A., Vandenberg, R. J., Arriza, J. L., Kavanaugh, M. P., and Amara, S. G. (1995). An excitatory amino-acid transporter with properties of a ligand-gated chloride channel. Nature 375, 599-603.
Hucka M, Finney A, Sauro H M, Bolouri H, Doyle J C, Kitano H et al. (2003) The Systems Biology Markup Language (SBML): A Medium for Representation and Exchange of Biochemical Network Models. Bioinformatics 9:524-531
Li C, Donizelli M, Rodriguez N, Dharuri H, Endler L, Chelliah V, Li L, He E, Henry A, Stefan MI, Snoep JL, Hucka M, Le Novere N, Laibe C (2010) BioModels Database: An enhanced, curated and annotated resource for published quantitative kinetic models. BMC Syst Biol., 4:92.
Otis, T., Kavanaugh, M., and Jahr, C. (1997). Postsynaptic glutamate transport at the climbing fiber-Purkinje cell synapse. Science 277, 1515.
Raikov I, De Schutter, E (2011) The layer-oriented approach to biological modeling languages. In revision.
Santamaria F, Wils S, De Schutter E, Augustine GJ (2006) Anomalous diffusion in Purkinje cell dendrites caused by dendritic spines. Neuron 52: 635-648.
Schmidt H, Stiefel KM, Racay P, Schwaller B, Eilers J (2003) Mutational analysis of dendritic Ca2+ kinetics in rodent Purkinje cells: role of parvalbumin and calbindin D28k. J Physiol 551:13-32.
Simoes de Souza F. M., De Schutter E. (2011) Robustness effect of gap junctions between Golgi cells on cerebellar cortex oscillations. Neural Systems & Circuits 1:7.
Slepoy A, Thompson AP, Plimpton SJ (2008) A constant-time kinetic Monte Carlo algorithm for simulation of large biochemical reaction networks. J Chem Phys 128: 205101.
Torben-Neilsen B, Uusisaari M, Stiefel KM (2010) A comparison of methods to determine neuronal phase-response curves. Front. Neuroinform. 4: 6.
Wils S, De Schutter E (2009) STEPS: Modeling and simulating complex reaction-diffusion systems with Python. Front Neuroinform 3: 15.
Zerangue, N., and Kavanaugh, M. P. (1996). Flux coupling in a neuronal glutamate transporter. Nature 383, 634-637.
4. Publications
4.1 Journals
- Uusisaari, M. & De Schutter, E. The mysterious microcircuitry of the cerebellar nuclei. Journal of Physiology (London), in press (2011).
- Tahon, K., Wijnants, M., De Schutter, E. & Maex, R. Current Source Density Correlates of Cerebellar Golgi and Purkinje Cell Responses to Tactile Input. J Neurophysiol 105, 1327-1341, doi:jn.00317.2010 [pii] 10.1152/jn.00317.2010 (2011).
- Steuber, V., Schultheiss, N., Silver, A., De Schutter, E. & Jaeger, D. Determinants of synaptic integration and heterogeneity in rebound firing explored with date-driven models of deep cerebellar nuclei cells. Journal of Computational Neuroscience, in press, doi:10.1007/s10827-010-0282-z (2011).
- Robberechts, Q., Wijnants, M., Giugliano, M. & De Schutter, E. Long-term depression at parallel fiber to Golgi cell synapses. J Neurophysiol 104, 3413-3423, doi:jn.00030.2010 [pii] 10.1152/jn.00030.2010 (2010).
- de Souza, F. & De Schutter, E. Robustness effect of gap junctions between Golgi cells on cerebellar cortex oscillations. Neural Systems & Circuits 1, 7 (2011).
- De Schutter, E. Data publishing and scientific journals: the future of the scientific paper in a world of shared data. Neuroinformatics 8, 151-153, doi:10.1007/s12021-010-9084-8 (2010).
- Chen, W., Maex, R., Adams, R., Steuber, V., Calcraft, L. & Davey, N. Clustering predicts memory performance in networks of spiking and non-spiking neurons. Frontiers in Computational Neuroscience, 2011.00014, doi:10.3389/fncom.2011.00014 (2011).
- Botta, P., de Souza, F. M., Sangrey, T., De Schutter, E. & Valenzuela, C. F. Alcohol excites cerebellar Golgi cells by inhibiting the Na+/K+ ATPase. Neuropsychopharmacology 35, 1984-1996, doi:npp201076 [pii] 10.1038/npp.2010.76 (2010).
- Anwar, H., Hong, S. & De Schutter, E. Controlling Ca2+-activated K+ channels with models of Ca2+ buffering in Purkinje cells Cerebellum, in press, doi:10.1007/s12311-010-0224-3 (2010).
4.2 Books and other one-time publications
Nothing to report.
4.3 Oral and Poster Presentations
- Warnaar, P. Properties of in vivo recorded Purkinje cell complex spikes, C7 Consortium meeting, Antwerp, Belgium, Jan 18. 2011, 2011
- Van Geit, W. & De Schutter, E. Automatically fitting a Purkinje cell model to experimental data using Neurofitter, FENS Forum, Amsterdam, the Netherlands, July 3-7. 2010, 2010
- Raikov, I. & De Schutter, E. A modeling language for large-scale neural network description, FENS Forum, Amsterdam, the Netherlands, July 3-7. 2010, 2010
- Publio, R. & De Schutter, E. The roles of ascending branch and parallel fiber synapse in cerebellar signaling, FENS Forum, Amsterdam, the Netherlands, July 3-7. 2010, 2010
- Negrello, M., Hong, S. & De Schutter, E. Complex spike related pauses in rat and monkey Purkinje neurons, FENS Forum, Amsterdam, the Netherlands, July 3-7. 2010, 2010
- Negrello, M., Hong, S. & De Schutter, E. What was the Purkinje doing while the monkey slept?, Computational Neuroscience, San Antonio, Texas, July 25-27, 2010, 2010
- Kim, Y., Sinclair, R. & De Schutter, E. Why are dendrites locally planar?, FENS Forum, Amsterdam, the Netherlands, July 3-7. 2010, 2010
- Hong, S., Van Geit, W., Anwar, H. & De Schutter, E. Computational modeling of cerebellar Purkinje neurons with complex and reduced morphologies, 40th Annual Meeting Society for Neuroscience, San Diego, Califorrnia, November 13-17, 2010, 2010
- Hepburn, I., Chen, W. & De Schutter, E. Reaction-Diffusion and Membrane Potential Simulation with STEPS, Neuroinformatics 2010, Kobe, Japan, August 30 - September 1. 2010, 2010
- Hepburn, I., Chen, W. & De Schutter, E. STEPS: reaction-diffusion and membrane potential simulation in complex 3d geometries, FENS Forum, Amsterdam, the Netherlands, July 3-7. 2010, 2010
- de Souza, F. M. & De Schutter, E. Robustness effect of gap junctions between Golgi cells on cerebellar oscillations, 40th Annual Meeting Society for Neuroscience, San Diego, Califorrnia, November 13-17, 2010, 2010
- de Souza, F. M. & De Schutter, E. Robustness effect of gap junctions between Golgi cells on cerebellar cortex oscillations, FENS Forum, Amsterdam, the Netherlands, July 3-7. 2010, 2010
- De Schutter, E. Using computational methods to investigate brain function at molecular, cellular and network levels, Korea Advanced Institute of Science & Technology (KAIST), Daejeon, Korea, Feb 16, 2011, 2011
- De Schutter, E. Bistability of the parallel fiber synapse is the basis of cerebellar LTD: a modeling study, Korea Institute for Science and Technology (KIST), Seoul, Korea, Feb, 17 2011, 2011
- De Schutter, E. Bistability of the Parallel Fiber Synapse is the Basis of Cerebellar LTD: a Modeling Study, PRESTO Workshop: Decoding and Control of Neural Information, Onna, Okinawa, Japan, November 5, 2010, 2011
- De Schutter, E. Stochastic modeling of signaling pathways involved in induction of cerebellar long-term depression, Experimental Biology 2011, Washington, DC, USA, April 9-13, 2011
- De Schutter, E. Bistability of the parallel fiber synapse is the basis of cerebellar LTD: a modeling study, University of Chicago, Chicago, USA, Nov 11, 2010, 2010
- De Schutter, E. New model description standards to facilitate multi-scale modeling, Neuroinformatics 2010, Kobe, Japan, August 30 - September 1. 2010, 2010
- De Schutter, E. Bistable behavior of the parallel fiber synapse is the basis of long-term depression: a stochastic modeling study, Computational Neuroscience Workshops, San Antonio, Texas, July 28-29, 2010, 2010
- De Schutter, E. Five challenging questions on DCN connectivity and function, The cerebellum: from neurons to higher control and cognition, Pavia, Italy, July 8-9, 2010, 2010
- De Schutter, E. Five challenging questions on DCN connectivity and function, FENS Satellite Event : Cerebellar Nuclei - Ins and Outs, Amsterdam, the Netherlands, July 2, 2010, 2010
- De Schutter, E. Computational modeling to study the effect of ethanol on cerebellar neurons, ISBRA 2010, Paris, France, September 13-16. 2010, 2010
- De Schutter, E. Stochastic modeling of LTD induction and AMPA receptor trafficking at the parallel fiber synapse, Neuro2010, Kobe, Japan, September 2-4. 2010, 2010
- Chen, W., Hepburn, I. & De Schutter, E. Tetrahedral mesh generation and visualization for stochastic reaction-diffusion simulation, Computational Neuroscience, San Antonio, Texas, July 25-27, 2010, 2010
- Anwar, H., Hong, S. & De Schutter, E. Generating dendritic Ca2+ spikes with different models of Ca2+ buffering in cerebellar Purkinje cells, FENS Forum, Amsterdam, the Netherlands, July 3-7. 2010, 2010
- Anwar, H., Hong, S. & De Schutter, E. Generating dendritic Ca2+ spikes with different models of Ca2+ buffering in cerebellar Purkinje cells, Computational Neuroscience, San Antonio, Texas, July 25-27, 2010, 2010
- Antunes, G. & De Schutter, E. Bistable induction of cerebellar long-term depression in a stochastic model, 40th Annual Meeting Society for Neuroscience, San Diego, Califorrnia, November 13-17, 2010, 2010
- Antunes, G. & De Schutter, E. Stochastic modeling of signaling pathways involved in induction of cerebellar long-term depression, US - JAPAN Brain Research Cooperative Program Workshop, Onna-son, Okinawa, October 4-6, 2010, 2010
- Antunes, G. & De Schutter, E. Stochastic model of the induction of cerebellar long-term depression, FENS Forum, Amsterdam, the Netherlands, July 3-7. 2010, 2010
5. Intellectual Property Rights and Other Specific Achievements
Nothing to report
6. Meetings and Events
6.1 Summer course
- Date: June 14 – July 1, 2010
- Venue: Seaside House, OIST
- Co-organizers : K. Doya, K. Stiefel, J. Wickens, OIST
- Co-sponsors : Japanese Neural Network Society
- Speakers:
- Amari, Shun-Ichi, RIKEN BSI, Japan
- Arbuthnott, Gordon, OIST, Japan
- Brette, Romain, École Normale Supérieure, France
- Dayan, Peter, University College London, UK
- De Schutter, Erik, OIST, Japan
- Doya, Kenji, OIST, Japan
- Gutkin, Boris, École Normale Supérieure, France
- Hellgren K., Jeanette, Karolinska Institute, Sweden
- Izhikevich, Eugene, Brain Corporation, USA
- Karpova, Alla, Janelia Farm, USA
- Kawato, Mitsuo, ATR, Japan
- Knöpfel, Thomas, RIKEN BSI, Japan
- Obermayer, Klaus, Bernstein Center Berlin, Germany Ohzawa, Izumi, Osaka University, Japan
- Sharpee, Tatyana, Salk Institute, USA
- Spruston, Nelson, Northwestern University, USA
- Stiefel, Klaus, OIST, Japan
6.2 Workshop
- Date: October 4-6, 2010
- Venue: Seaside House, OIST
- Co-organizers:
- Lindsey B., Univeristy South Florida, USA
- Schwaber, James, Thomas Jefferson University, USA
- Co-sponsors: NIH, USA and NIPS, Japan
- Speakers:
- Antunes Gabriela, OIST, Japan
- Arata, Akiko , Hyoga College of Medicine, Japan
- Dick, Thomas E., Case Western Reserve Univ., USA Freeman, Kate, Thomas Jefferson University, USA
- Gumus, Zeynep, Cornell University, USA
- Hong, Kyonsoo, NewYork University, USA Ishii, Shin, Kyoto University, Japan
- Ito, Masao, RIKEN BSI, Japan
- Kuroda, Shinya, University of Tokyo, Japan
- Kuwaki, Tomoyuki, Kagoshima University, Japan
- Lindsey Bruce Univeristy South Florida, USA
- Morris, Kendall, University of South Florida, USA
- Noma, Akinori, Kyoto University, Japan
- Oku, Yoshitaka, Hyogo College of Medicine, Japan
- Pitts, Teresa, University of Florida, USA
- Rybak, Ilya, Drexel University, USA
- Schwaber, James, Thomas Jefferson University, USA
- Toporikova, Natalia,Georgia Institute Technology, USA
- Ueda, Hiroki R., RIKEN, Japan
- Zucker, Robert S., U C Berkeley, USA