



危険な毒蛇ハブの全ゲノム解読〜毒を作り出す遺伝子進化の全貌を世界で初めて解明〜

ハブ(Protobothrops flavoviridis)は国内の毒蛇としてよく知られており、その毒液は多様な生理活性を持つタンパク質の「カクテル」です。その全容解明のために、全ゲノム解読が待たれていました。九州大学生体防御医学研究所の柴田弘紀准教授は、沖縄科学技術大学院大学の佐藤矩行教授、東北大学の小川智久准教授らとの共同研究で、ハブの全ゲノム配列を決定し、ハブゲノムにコードされる約25,000個の遺伝子を発見しました。さらに毒液の成分として働くタンパク質の遺伝子60個と、それらと兄弟のタンパク質でありながら毒として働かない遺伝子(非毒型パラログ)を224個見出しました。毒液関連遺伝子のうち、特に4つのタンパク質ファミリー(金属プロテアーゼ、ホスホリパーゼA2、セリンプロテアーゼ、C型レクチン)では、遺伝子のコピー数が大幅に増加し、かつコピー間のアミノ酸の置換速度が上昇していること(加速進化)がわかりました。また、毒液関連遺伝子群が、鳥類や爬虫類に特徴的な組み替え率が高い小型の染色体、「微小染色体」に多く存在していることも見出しました。これらのことから、ハブ毒液遺伝子群が、高度に多重化かつ急速に多様化しながら進化してきたことが示唆されました。本成果により、蛇毒の作用機序の全容解明と、効果の高い抗毒素開発の大幅な効率化、さらにハブゲノム由来の新規の薬理分子からの有用な医薬品開発への道が開かれました。
本研究成果は国際学術誌 Scientific Reports に掲載されました。
なお本研究は、日本学術振興会科学研究費(JP25440214,JP18H02498,JP23107505,JP24651130)の支援を受けて行われました。
研究の背景
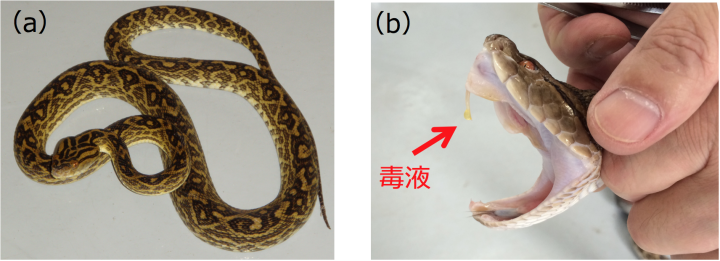
ハブは南西諸島に生息し、国内産のヘビの中では最も恐れられている毒蛇です(図1)。比較的大型のために攻撃射程が長く、注入される毒量も多いため、咬症被害が多く、特定動物に指定されています。多くの生息地では、衛生動物として駆除対象とされ、ハブ酒やハブ皮革製品などの産業利用も盛んです。その一方で、ハブの生息地である南西諸島には大型肉食動物が分布しないため、生態系において非常に重要な位置を占める捕食者でもあります。ハブをはじめとするクサリヘビ類の毒液は、出血毒として知られています。毒液は、血管を破壊する金属プロテアーゼ、炎症や壊死を引き起こすホスホリパーゼA2、血液を固まらせないC型レクチンなど、多様な生理活性を持つタンパク質の「カクテル」であり、その全容解明のためには、全ゲノム解読が待たれていました。
これまでヘビ類で全ゲノム配列が報告されていたのは、ビルマニシキヘビ、キングコブラ、ヒャッポダ、タイワンハブの4種でしたが、いずれも遺伝子カタログ作成に止まっており、遺伝子と染色体との関連までを解析対象とする真の「ゲノム解析」がなされたものはありませんでした。
研究の内容
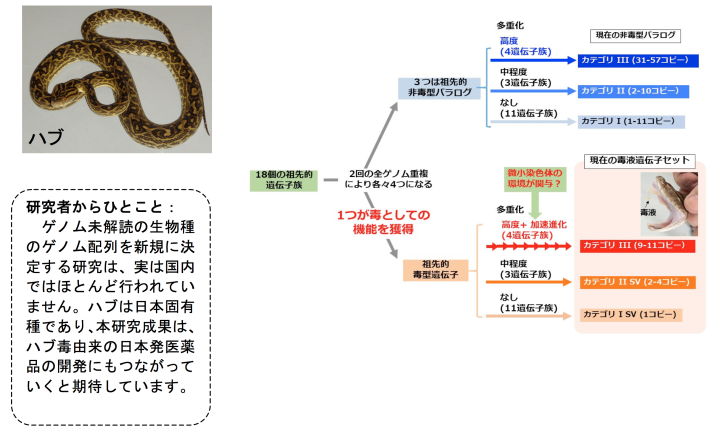
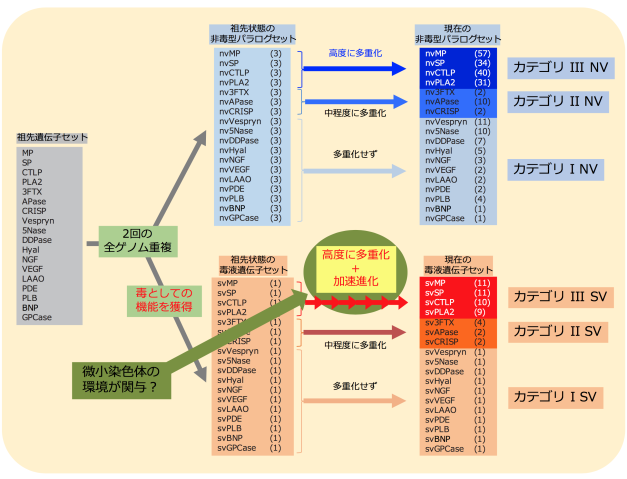
奄美大島産のハブからゲノムDNAを抽出し、超並列シークエンサで解析し、10億本のDNA断片(合計で136 Gb)データを取得しました。これらをつなぎあわせ、全長1.4 Gbのハブゲノムドラフト配列HabAm1として繋ぎあわせることができました。また、ハブの18種類の臓器・組織からRNAを抽出し、超並列シークエンサで配列を決定し、各組織において発現している遺伝子の情報としました。この情報を利用して、HabAm1から、25,134個の遺伝子を見つけることができました。この中から、毒液タンパク質遺伝子を60個および、非毒型パラログ(註1)を224個見つけることができました。これらは18個の遺伝子族に分類でき、遺伝子重複の度合いによっておおむね以下の3つのカテゴリに分類することができました(表1)。カテゴリIIIは、毒液の主要構成成分である金属プロテアーゼ(MP、註2)、セリンプロテアーゼ(SP、註3)、C型レクチン(CTLP、註4)、ホスホリパーゼA2(PLA2、註5)の4遺伝子族からなり、毒液遺伝子コピーと非毒型パラログともに高度に多重化していました。また、三本指型トキシン(3FTX、註6)、アミノペプチダーゼ(APase、註7)、システインリッチ分泌タンパク質(CRISP、註8)の3族は毒液遺伝子コピー、非毒型パラログのいずれも中程度の多重化を示していました(表1中のカテゴリII)。それ以外の遺伝子族では毒型遺伝子1コピーと非毒型パラログ2〜10コピーであり(表1中のカテゴリI)、 脊椎動物の初期進化過程で起きたとされる2回のゲノム重複によってできた4つの遺伝子コピーの中の1コピーだけが毒液機能を獲得したことが示唆されました。
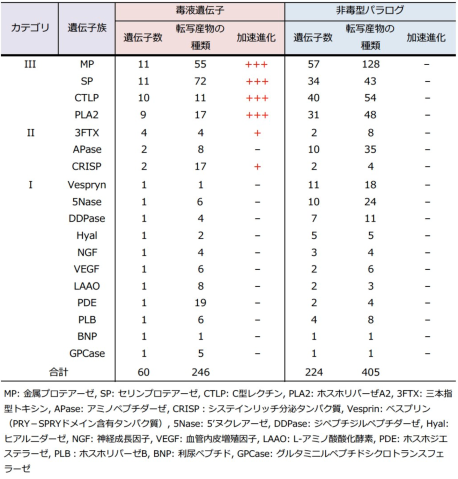
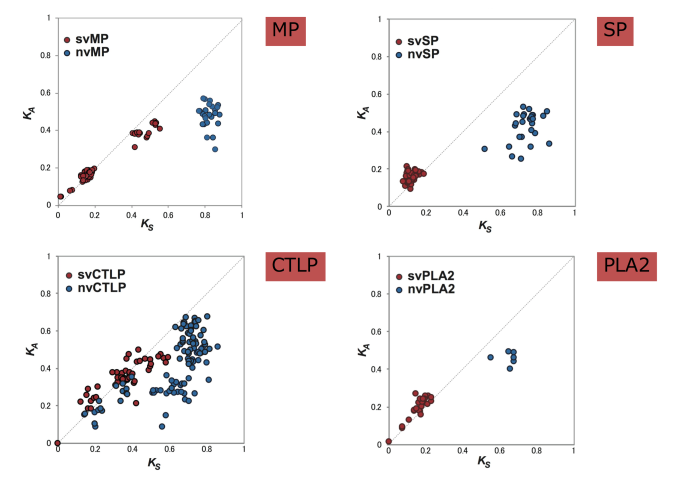
また、毒液関連遺伝子の進化速度(厳密にはコピー間の分化の速度)を比較したところ、カテゴリIの毒液タンパク質遺伝子群においてのみ顕著な加速進化(註9)が観察されました(図2)。それに対してそれぞれの非毒型パラログ群では加速進化の傾向は全く観察されませんでした。
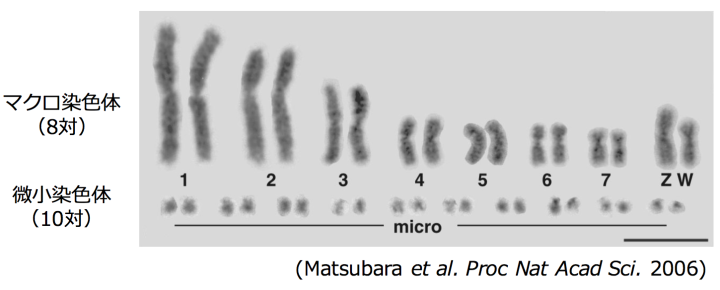
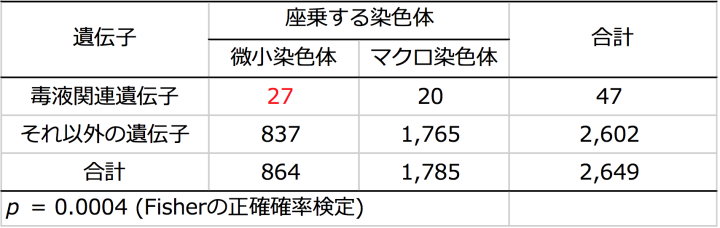
さらに、ヘビの中で最もくわしく染色体が調べられているシマヘビとの比較で、ハブゲノム上の2,649個の遺伝子について座乗する染色体を決定できました。染色体が特定できた遺伝子を毒液関連遺伝子群とそれ以外の遺伝子群とに分類すると、毒液関連遺伝子群が微小染色体(図3、註10)に濃縮していることがわかり、この偏りは統計学的に有意でした(p < 0.0004, Fisherの正確確率検定)(次頁の表2)。このことは、毒液タンパク質遺伝子群において観察された加速進化には、微小染色体の特異なゲノム環境が関与していると考えられました(下図4)。
今後の展開
今回のゲノム解読から得られた毒液タンパク質のうち機能の詳細が不明のものがまだたくさんあります。たとえば、これまでコブラ類(主に神経毒を産生する)からのみ知られていた三本指型トキシンも、ハブのゲノム中に存在することがわかりました。これらの機能解析を進めていくことで、毒液の作用機序の完全解明に大きく近づくことになります。またハブは特定動物に指定されており、多くの地域では駆除対象となっています。しかし上述のように生息地では重要な捕食者として生態系のKeystone種でもあります。本研究で得られたゲノム配列を基礎情報として、ハブの自然集団における遺伝的多様性の研究が可能になり、これまでほとんど省みられることのなかったハブの種の保全についての科学的検証が可能となりました。
用語解説
注1:非毒型パラログ
遺伝子重複によって生じた遺伝子コピーのそれぞれをパラログといい、そのうち毒としての機能をもっていないタンパク質の遺伝子を指す。
注2:金属プロテアーゼ(MP)
タンパク質分解酵素。組織基底膜を壊して出血を引き起こしたり,細胞死を誘導したりする機能を持つ。ハブ毒液の主要な成分の一つ。
注3:セリンプロテアーゼ(SP)
タンパク質分解酵素。フィブリノーゲンや血液凝固系因子に特異的に作用し、血液凝固を部分的に引き起こす。血管内での微小な凝固を引き起こすことで結果的にフィブリノーゲンを消費させて出血を起こす。ハブ毒液の主要な成分の一つ。
注4:ホスホリパーゼA2(PLA2)
リン脂質を分解する酵素。赤血球の細胞膜を壊して溶血を引き起こしたり、腫れや神経毒性、筋肉の壊死など、多様な病理的症状を引き起こすハブ毒液の主要な成分の一つ。
注5:C型レクチン(CTLP)
Ca2+イオン依存的に糖鎖に特異的に結合するレクチン。ハブ毒中には,レクチンとしての機能を持つもののほか,血液凝固第IX, 第X因子などに作用して血液凝固を阻害したり,血小板の凝集・活性化や細胞接着に関わるインテグリン分子に作用して阻害する。ハブ毒液の主要な成分の一つ。
注6:三本指型トキシン(3FTX)
コブラやウミヘビなどの神経毒の主成分として知られ,アセチルコリン受容体や様々なチャネルタンパク質に特異的に作用する。ハブに見出された三本指型トキシンの機能については不明である。
注7:アミノペプチダーゼ(Apase)
タンパク質、ペプチドのアミノ末端側から順次アミノ酸を遊離する酵素。ハブ毒中のペプチド毒成分の成熟化、活性化に働く。
注8:システインリッチ分泌タンパク質(CRISP)
16個のシステインが保存されたタンパク質ファミリー。ヒトでは精巣に発現し授精などに関与している。ヘビ毒CRISPは,Kチャネルやリアノジン受容体などのイオンチャネルをブロックする。
注9:加速進化
通常のタンパク質には機能的制約があるため、分子進化学的解析においてはアミノ酸を変える塩基置換(非同義置換)よりも、アミノ酸を変えない塩基置換(同義置換)の方が多く観察される。しかし一部の遺伝子では、非同義置換が同義置換と同等またはそれ以上多く観察されることがあり、通常よりも早い速度でアミノ酸置換が起こっているという意味で「加速進化」と呼ぶ。
注10:微小染色体
ヘビを含む爬虫類や鳥類の染色体はヒトと同様な大型の染色体と小型の染色体(微小染色体)の2種類から成り立っており、大型の染色体をマクロ染色体、小型の染色体を微小染色体と呼ぶ(図3)。ニワトリの研究から、微小染色体は、マクロ染色体に比較して、遺伝子に富むこと、GC含有率が高いこと、組み換え率が高いことなどが知られている。
---
共同プレスリリース(PDF)











